TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025016632
公報種別
公開特許公報(A)
公開日
2025-02-04
出願番号
2024191089,2021548555
出願日
2024-10-30,2019-10-30
発明の名称
GRAMC:シス調節モジュールのゲノムスケールレポーターアッセイ法
出願人
ラトガーズ、ザ ステイト ユニバーシティ オブ ニュージャージー
代理人
個人
,
個人
,
個人
,
個人
,
個人
主分類
C12Q
1/6806 20180101AFI20250128BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】GRAMC:シス調節モジュールのゲノムスケールレポーターアッセイ法の提供。
【解決手段】本明細書には、機能的調節エレメントのレポーター核酸のライブラリー、ならびにそのようなライブラリーを構築および使用するための方法およびキットが開示されている。例示的なライブラリー、方法、およびキットは、機能的核酸調節エレメントのハイスループット検出、同定、および/または定量化のために使用することができる。一部の例では、核酸は、目的の細胞または目的の細胞の集団から得られるゲノムDNAである。ゲノムDNAは、これらに限定されないが、動物(例えば、哺乳動物)、植物、細菌、真菌、または古細菌を含む、任意の目的の生物に由来してもよい。
【選択図】なし
特許請求の範囲
【請求項1】
明細書に記載の発明。
発明の詳細な説明
【技術分野】
【0001】
関連出願の相互参照
本出願は、参照によりその全体が本明細書に組み込まれる、2018年10月31日に出願された米国仮出願第62/753,608号の利益を主張する。
続きを表示(約 4,000 文字)
【0002】
分野
本出願は、レポーター核酸、例えば機能的調節エレメントのライブラリー、ならびにそのようなライブラリーを構築および使用するための方法およびキットを提供する。
【背景技術】
【0003】
エンハンサー、プロモーター、およびリプレッサーなどのシス調節モジュール(CRM)は、ゲノムの機能的エレメントである。ヒトゲノムにわたって数十万個のCRMが散在していると推定されている(Niu, et al. Nucleic acids research 46.11 (2018): 5395-5409; Visel, et al. Nature 461.7261 (2009):199;ENCODE Project Consortium. Nature 489.7414 (2012):57)。CRMは、遺伝子が、いつ、どこで、ど
のレベルで発現されるかを調節するため、CRMは、ほぼすべての生物学的プロセスに関与する。個々のCRMが複数の転写因子と直接的に相互作用し、複数のCRMが一緒になって機能して遺伝子調節活性を媒介する(Davidson. The Regulatory Genome, Elsevier (2006); Levine, et al. Cell 157.1 (2014): 13-25;De Laat, et al.
Nature 502.7472 (2013): 499)。こうしたエレメントの包括的な実験的同定は困難である。
【0004】
CRMを同定するための標準的なレポーターアッセイは、基本プロモーターおよびレポーター遺伝子の上流にある候補CRMをクローニングし、レポーター遺伝子の発現を駆動するその能力を調査することである(Rosenthal, Methods in enzymology 152 (1987): 704-720;Arnone, et al. Methods in cell biology 74. (2004): 621-652;Banerji, et al. Cell 27.2 (1981): 299-308)。同じレポーター構築物により、CRMが、遺伝子摂動に対して(Nam, et al.PLoS One 7.4 (2012): e35934.)
、および転写結合部位の変異に対して(Damle, et al. Developmental biology 357.2 (2011): 505-517;de-Leon, et al. PNAS USA 107.22 (2010): 10103-10108;Cui, et al. Cell reports 19.2 (2017): 364-374;Emison, et al. Nature
434.7035 (2005): 857;Guerreiro, et al. PNAS USA 110.26 (2013): 10682-10686)、どのように応答するかをモニターすることができる。しかしながら、このような従来の1つずつのレポーターアッセイは、ゲノムに含有されている何百万個もの潜在的なCRMの分析(例えば、ハイスループット分析)には好適ではない。幾つかのハイスループットアッセイが試みられているが、バイアスが問題となることがある。
【先行技術文献】
【非特許文献】
【0005】
Rosenthal、Methods in enzymology(1987)152:704~720
Arnoneら、Methods in cell biology(2004)74:621~652
Banerjiら、Cell(1981)27.2:299~308
【発明の概要】
【課題を解決するための手段】
【0006】
本明細書には、核酸分子レポーターライブラリーを構築する方法、および本明細書に開示されている方法を使用して産生される核酸分子レポーターライブラリーが開示されている。本開示のゲノムスケールレポーターアッセイ法は、標準的なレポーターアッセイの場合と同様に、エンハンサーおよびプロモーターの両方に対して効果的である。本アッセイは、長鎖DNAインサートにも対応し、部分的なCRMではなく完全なCRMのスクリーニングを可能にする。ゲノムカバレッジおよびDNAバーコードが過剰であると実験コストが増加し、ゲノムカバレッジおよびDNAバーコードが不十分であると、信頼性の低いデータがもたらされる。しかしながら、本明細書で開示されているライブラリーおよび方法では、ゲノムカバレッジおよびライブラリー内のDNAバーコードの数は調整可能である。最後に、本アッセイは、現在利用可能な方法と同等のまたはそれよりも少ない入力材料で、再現性のあるデータを生成する。
【0007】
一部の実施形態では、核酸分子レポーターライブラリーを構築する方法は、選択されたサイズ範囲(例えば、約750~850塩基対長など、100~3000塩基対長のサイズ範囲)の複数の核酸分子(例えば、ゲノムDNAまたは合成DNA)を単離するステップ;複数の単離された核酸分子を、少なくとも1つの線状アダプター配列(3’末端の少なくとも1つのデオキシリボヌクレオチドおよび5’末端の少なくとも1つのデオキシリボヌクレオチドが隣接する少なくとも2つの連続したリボヌクレオチドを含むアダプターなど)にライゲートして、インサート(単離された核酸分子)およびアダプターを含む複数の環状核酸分子を形成するステップ;複数の環状核酸分子を、複数の線状核酸分子を産生するのに十分な条件下で酵素と接触させるステップ;ならびに複数の線状核酸分子を少なくとも1つのレポーター核酸と融合させて複数のレポーター構築物を産生し、核酸分子レポーターライブラリーを形成するステップを含む。
【0008】
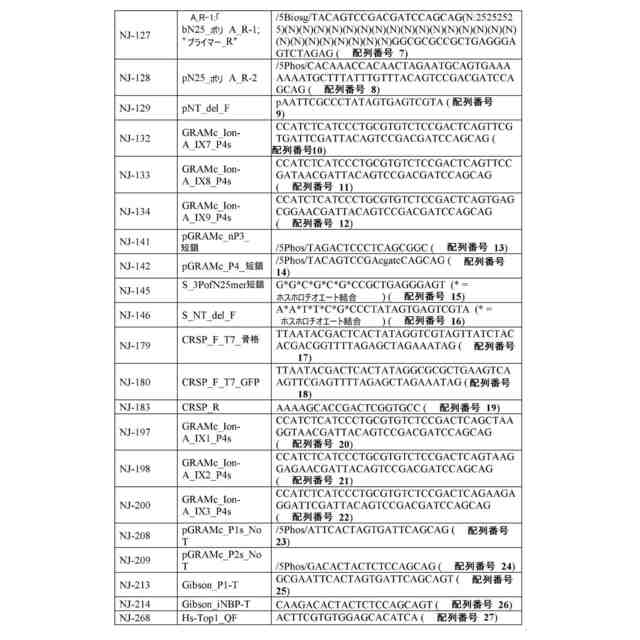
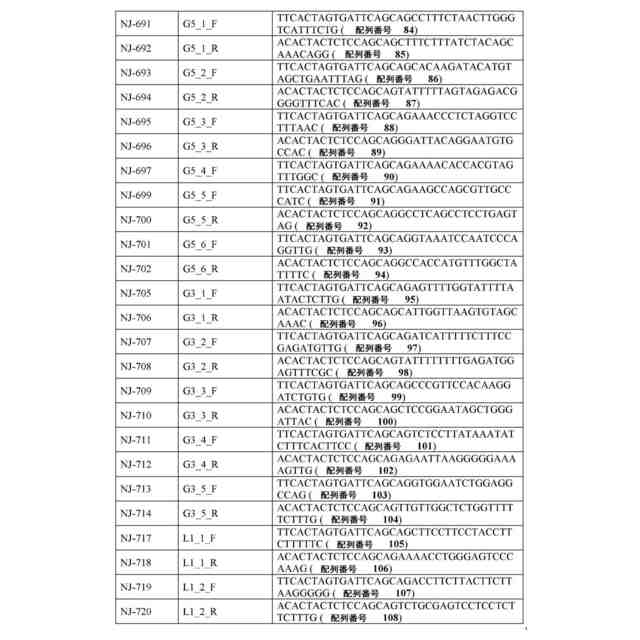
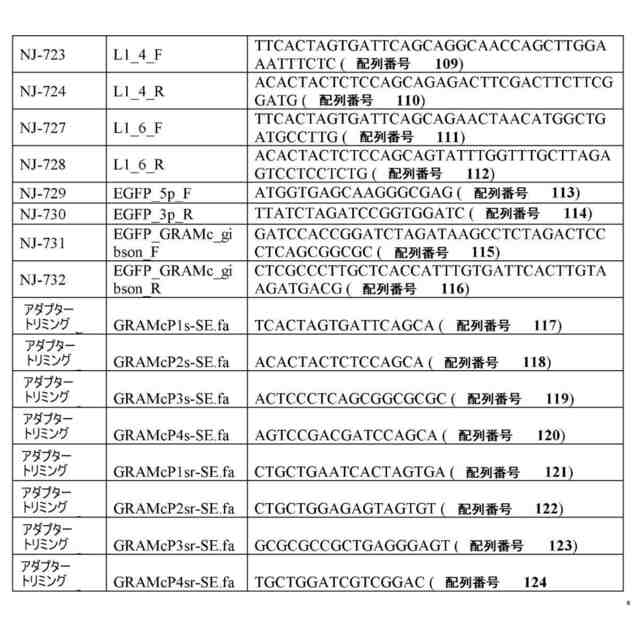
ゲノムDNA(ゲノムDNA断片など)または合成DNAを含む、任意の核酸分子を使用することができる。一部の例では、核酸は、目的の細胞または目的の細胞の集団から得られるゲノムDNAである。ゲノムDNAは、これらに限定されないが、動物(例えば、哺乳動物)、植物、細菌、真菌、または古細菌を含む、任意の目的の生物に由来してもよい。一部の例では、本方法は、ゲル電気泳動またはビーズに基づくサイズ選択を使用して、単離された核酸分子のサイズ範囲を選択するステップを含む。一部の例では、本方法は、複数の単離された核酸分子を、リガーゼを使用して少なくとも1つの線状アダプター配列にライゲートするステップを含む。一部の例では、リガーゼは、T4 DNAリガーゼなどのDNAリガーゼを含む。線状アダプター配列は、3’末端の少なくとも1つのデオキシリボヌクレオチドおよび5’末端の少なくとも1つのデオキシリボヌクレオチドが隣接する少なくとも2つの連続したリボヌクレオチド(例えば、配列番号1および/または配列番号2の核酸)を含んでいてもよい。したがって、ライゲーションにより、インサートおよびアダプターを含む複数の環状核酸分子が産生される。
【0009】
一部の例では、本方法は、環状核酸を線状化する前に、複数の環状核酸分子を、複数の環状核酸分子から線状核酸分子を除去するのに十分な条件下で、エキソヌクレアーゼ(例えば、エキソヌクレアーゼI、エキソヌクレアーゼIII、および/またはラムダエキソヌクレアーゼ)と接触させるステップをさらに含む。一部の例では、本方法は、次いで、複数の環状核酸分子を、インサートが隣接する、各々が3’末端の少なくとも1つのデオキシリボヌクレオチドおよび5’末端の少なくとも1つのデオキシリボヌクレオチドを含む複数の線状核酸分子を産生するのに十分な条件下で、エンドリボヌクレアーゼ(例えば、RNase HIIまたはウラシル-DNAグリコシラーゼなどの、DNA二重鎖内のリボヌクレオチドに特異的なエンドリボヌクレアーゼ)と接触させるステップを含む。一部の例では、本方法は、複数の線状核酸分子を少なくとも1つのレポーター核酸(例えば、蛍光タンパク質をコードする核酸および/またはバーコードを含む核酸)と融合させて、複数のレポーター構築物を産生するステップを含む。
【0010】
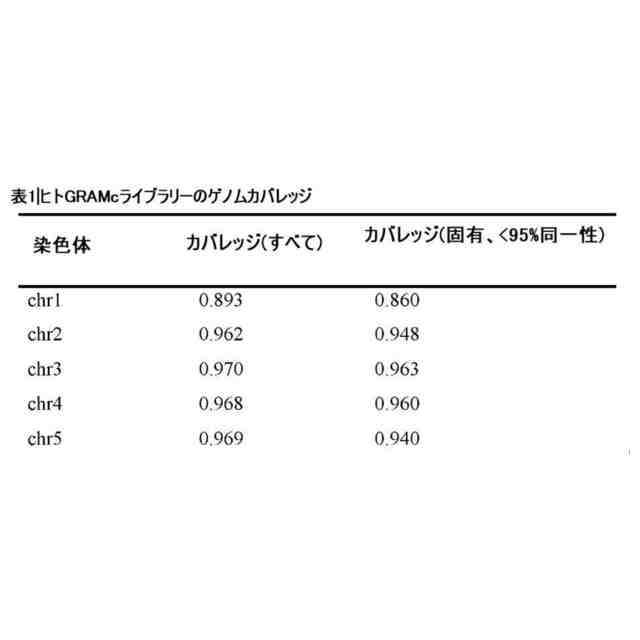
一部の例では、本方法は、複数の線状核酸分子のゲノムカバレッジを決定するステップをさらに含む。例えば、ゲノムカバレッジを決定するステップは、少なくとも1つの目的のゲノム領域を選択するステップ、複数の線状核酸分子を増幅するステップ、ならびに選択されたゲノム領域が複数の線状核酸分子に存在するか否か、複数の線状核酸分子における選択されたゲノム領域のコピー数、および/またはゲノムカバレッジを決定するステップを含んでいてもよい。一部の例では、ゲノムカバレッジは、分析のために1つまたは複数の単一コピー標的を選択することにより決定される。例示的な単一コピー標的としては、ACTA1、ADM、ADAM12、AXL、CFB、DLX5、Kiss1、NCOA6、Notch2、RPP30、およびTOP1が挙げられる。ライブラリーの出発材料の供給源に応じて、追加のまたは代替の単一コピー標的を選択することができる。
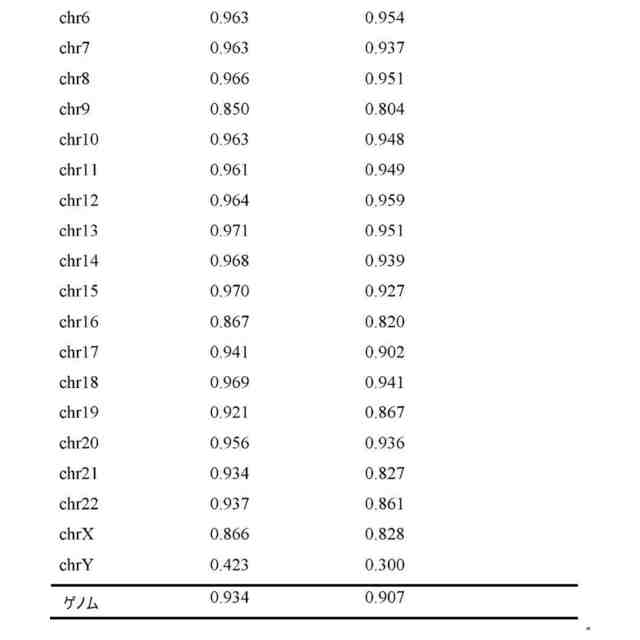
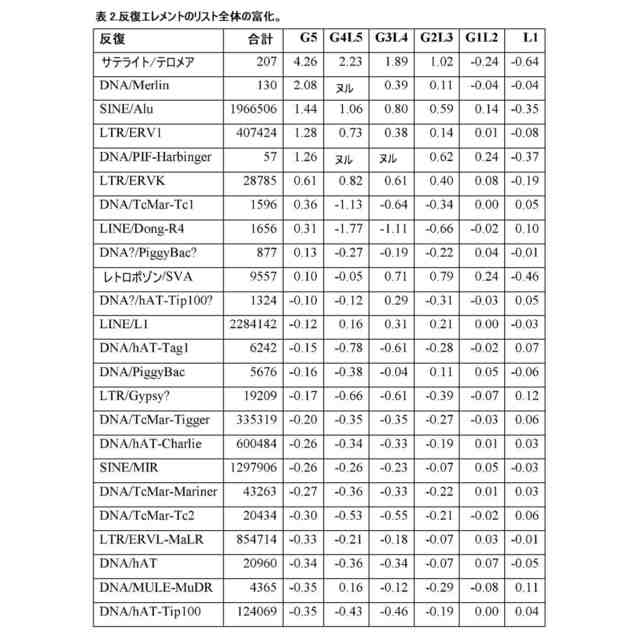
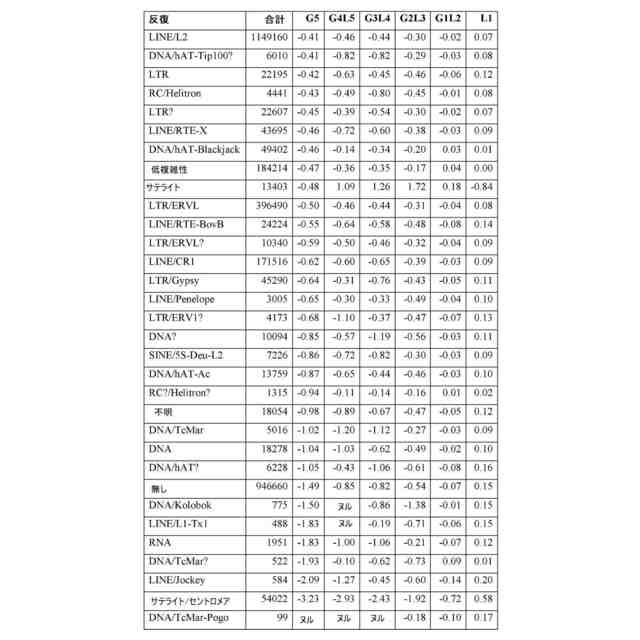
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
個人
酒類
2か月前
株式会社オシキリ
発酵検査装置
2か月前
株式会社ゴーフォトン
PCR方法
7日前
個人
セルロース性物質の製造方法
1か月前
松谷化学工業株式会社
澱粉分解物の製造方法
2か月前
SMC株式会社
気体供給装置
17日前
東ソー株式会社
ポリマーコートビーズの製造方法
2か月前
株式会社GSP研究所
miRNA検出方法
1か月前
鹿島建設株式会社
褐藻の冷凍保存方法
3日前
サッポロビール株式会社
アルコール飲料
1か月前
サッポロビール株式会社
アルコール飲料
1か月前
サッポロビール株式会社
アルコール飲料
3日前
株式会社テクノーブル
乳酸菌及び皮膚外用剤
3日前
アサヒビール株式会社
容器詰麦芽発酵飲料
1か月前
新東工業株式会社
培養システム
11日前
株式会社SUPER BLOOM
スピリッツ
3日前
新東工業株式会社
培養システム
11日前
東ソー株式会社
アデノ随伴ウイルスのスクリーニング方法
2か月前
株式会社エムスタイル
微生物群の賦活化方法
1か月前
学校法人君が淵学園
核酸の部位特異的アシル修飾剤
1か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
1か月前
三井化学株式会社
培養容器
2か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
1か月前
サッポロビール株式会社
茶風味アルコール飲料
19日前
ヤマサ醤油株式会社
アルギニン非分解性乳酸菌株の取得方法
24日前
株式会社渡辺オイスター研究所
脂肪細胞の成長抑制剤
2か月前
学校法人近畿大学
好気性微生物の培養方法および培養器
1か月前
サムコ株式会社
ポアデバイス
1か月前
国立大学法人富山大学
T細胞受容体およびその利用
1か月前
学校法人帝京大学
油脂を過剰蓄積する油脂生産酵母変異株
3日前
株式会社豊田中央研究所
環境核酸モニタリング方法
1か月前
雪印メグミルク株式会社
D-セリン濃度低減用組成物
2か月前
株式会社トクヤマ
乾燥用保護剤、生体試料測定試薬、及び分析方法
2か月前
公立大学法人宮城大学
D-セリンの製造方法
2か月前
ウシオ電機株式会社
培養容器
2か月前
ZACROS株式会社
培養装置、及び培養装置積層体
2か月前
続きを見る
他の特許を見る


 特許ウォッチ
特許ウォッチ