TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2024087054
公報種別
公開特許公報(A)
公開日
2024-06-28
出願番号
2024071857,2021073812
出願日
2024-04-25,2016-07-04
発明の名称
微生物油産生ラビリンチュラ類、微生物油、ならびにそれらの作成方法およびそれらの使用
出願人
国立大学法人九州大学
,
学校法人甲南学園
,
株式会社ニッスイ
代理人
個人
,
個人
主分類
C12P
7/6427 20220101AFI20240621BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】 エロンガーゼ/デサチュラーゼ経路のみでPUFAを生産するラビリンチュラを提供すること。
【解決手段】 内因性PUFA-PKS経路によるPUFAの生産能が無いあるいは極めて微弱であり、かつ内因性エロンガーゼ/デサチュラーゼ経路によるPUFA生産能を有するラビリンチュラ類。パリエティキトリウム属(
Parietichytrium
)またはシゾキトリウム属(
Schizochytrium
)のいずれかに属する微生物である。
【選択図】 図1
特許請求の範囲
【請求項1】
脂肪酸組成が改変するように遺伝子改変されたラビリンチュラ類から、微生物油を製造する方法であって、ラビリンチュラ類が、内因性PUFA-PKS経路による高度不飽和脂肪酸(PUFA)の生産能が無いあるいは極めて微弱であり、かつ内因性エロンガーゼ/デサチュラーゼ経路によるPUFA生産能を有するParietichytrium属またはSchizochytrium属に属し、ラビリンチュラ類の脂肪酸不飽和化酵素遺伝子を破壊または/および発現抑制することにより、脂肪酸組成が改変されたラビリンチュラ類であって、該ラビリンチュラ類から得られる微生物油が、下記(a)および(b)を満たす微生物油であることを特徴とする、微生物油の製造方法。
(a)DHAが全脂肪酸組成のうち、0.50%以下である微生物油。
(b)DHAとn-6DPAの合計が全脂肪酸組成のうち、0.7%以下である微生物油。
続きを表示(約 1,300 文字)
【請求項2】
ラビリンチュラ類の遺伝子を破壊する方法が、エレクトロポレーション、遺伝子銃法またはゲノム編集である、請求項1に記載の微生物油の製造方法。
【請求項3】
ラビリンチュラ類の遺伝子を発現抑制する方法が、アンチセンス法あるいはRNA干渉である、請求項1に記載の微生物油の製造方法。
【請求項4】
脂肪酸不飽和化酵素遺伝子がΔ4デサチュラーゼ遺伝子である、請求項1ないし3のいずれかに記載の微生物油の製造方法。
【請求項5】
Parietichytrium属に属するラビリンチュラ類が、Parietichytrium sarkarianumに属し、Schizochytrium属に属するラビリンチュラ類が、Schizochytrium aggregatumに属するラビリンチュラ類である、請求項1ないし4のいずれかに記載の微生物油の製造方法。
【請求項6】
Parietichytrium属に属するラビリンチュラ類が、Parietichytrium sp.SEK358、FERM BP-11405、Parietichytrium sarkarianum SEK364、FERM BP-11298またはParietichytrium sp.SEK571、FERM BP-11406であり、Schizochytrium属に属するラビリンチュラ類が、Schizochytrium aggregatum ATCC2829である、請求項1ないし4のいずれかに記載の微生物油の製造方法。
【請求項7】
ラビリンチュラ類の脂肪酸組成の改変方法であって、ラビリンチュラ類が、内因性PUFA-PKS経路による高度不飽和脂肪酸(PUFA)の生産能が無いあるいは極めて微弱であり、かつ内因性エロンガーゼ/デサチュラーゼ経路によるPUFA生産能を有するParietichytrium属またはSchizochytrium属に属し、ラビリンチュラ類の脂肪酸不飽和化酵素遺伝子を破壊または/および発現抑制することにより、脂肪酸組成が改変されたラビリンチュラ類であって、該ラビリンチュラ類から得られる微生物油が、下記(a)および(b)を満たす微生物油であることを特徴とする、脂肪酸組成の改変方法。
(a)DHAが全脂肪酸組成のうち、0.50%以下である微生物油。
(b)DHAとn-6DPAの合計が全脂肪酸組成のうち、0.7%以下である微生物油。
【請求項8】
ラビリンチュラ類の遺伝子を破壊する方法を、エレクトロポレーション、遺伝子銃法またはゲノム編集によって行う、請求項7に記載の脂肪酸組成の改変方法。
【請求項9】
ラビリンチュラ類の遺伝子を発現抑制する方法が、アンチセンス法あるいはRNA干渉である、請求項7に記載の脂肪酸組成の改変方法。
【請求項10】
脂肪酸不飽和化酵素遺伝子がΔ4デサチュラーゼ遺伝子である、請求項7ないし9のいずれかに記載の脂肪酸組成の改変方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、ラビリンチュラ類から得られる微生物油、微生物油産生ラビリンチュラ類、ならびに微生物油産生ラビリンチュラ類の使用に関する。
より詳細には、高度不飽和脂肪酸(PUFA)、その生産に関するものであり、
脂肪酸組成が改変するように遺伝子改変されたラビリンチュラ類、好ましくはエロンガーゼ/デサチュラーゼ経路のみでPUFAを生産するラビリンチュラ類、それらを用いたPUFAの製造方法およびそれらを用いて製造されたPUFAを含有する脂質(微生物油)およびそれらの使用に関する。
続きを表示(約 5,200 文字)
【背景技術】
【0002】
ラビリンチュラ類(
Labyrinthulids
)はラビリンチュラ目(Order
Labyrinthulales
)、ヤブレツボカビ目(Order
Thraustochytriales
)を含むラビリンチュラ綱(Class
Labyrinthulomycetes
)に属する真核微生物で、海洋に普遍的に存在することが知られている。また、ヤブレツボカビ目に属する微生物はヤブレツボカビ類(
Thraustochytrids
)と総称されることもある。
ラビリンチュラ類は、産業的には油糧微生物として注目されており、これらラビリンチュラ類が生産するDHAはDHA含有脂質の原料や、高DHA含有飼料等が製品化されている(非特許文献1)。具体的には、例えば、トラウストキトリウム属、シゾキトリウム属の生育技術(特許文献1)、トロウストチトリアレ類、トロウストチトリアレ類から抽出されるω-3HUFA(高度不飽和脂肪酸)の利用技術(特許文献2)等が挙げられる。
【0003】
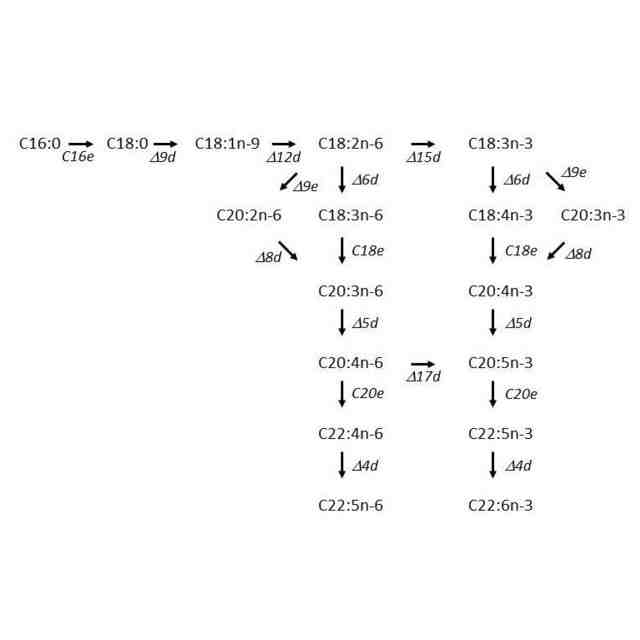
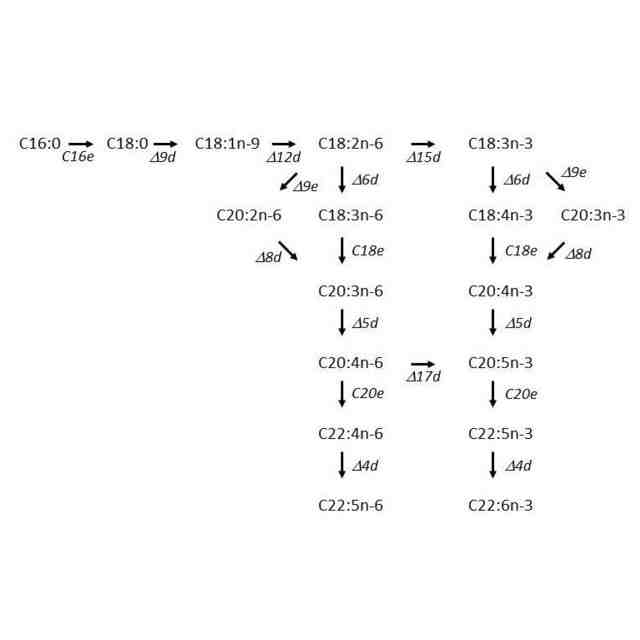
一般に、PUFAはエロンガーゼ/デサチュラーゼ経路(スタンダード経路と呼ばれることもある。)により生合成されることが知られているが、ある種のラビリンチュラ類はこれとは異なる経路、すなわちポリケチド合成酵素(ポリケチドシンターゼ、polyketide synthase、PKS)による代謝経路でPUFAを生産することが明らかにされている(非特許文献2)。以下、本発明ではこの経路をPUFA-PKS経路またはPKS経路と呼ぶ。この経路によって生産されるPUFAの組成は、そのほとんどがDHAとDPAn-6という特徴を有する。
【0004】
ラビリンチュラ類(特にヤブレツボカビ類)はPUFA生合成経路としてPUFA-PKS経路のみを有し、他の生物では一般的なエロンガーゼ/デサチュラーゼ経路を有さないと考える科学者もいる(非特許文献3)。実際、ラビリンチュラ類のPUFA-PKS経路の遺伝子破壊により当該ラビリンチュラ類は致死性となり、培地中にPUFAを添加しなければ生育できなくなったとの報告を例示することができる(非特許文献3)。この結果は、当該ラビリンチュラ類の生育にはPUFAが必須であり、唯一のPUFA生合成経路であるPUFA-PKS経路の遺伝子が破壊されたため、生育のために外因性のPUFAを要求する性質に変化したことを意味する。
【0005】
しかしこのような当業者の常識に反して、我々は検討の結果、PUFA生合成経路としてPUFA-PKS経路とエロンガーゼ/デサチュラーゼ経路の双方を有するラビリンチュラ類が存在することを見出した。具体的には特許文献3の明細書や非特許文献5に詳述されている。
Thraustochytrium
aureum
ATCC34304を例示すると、この株がエロンガーゼ/デサチュラーゼ経路のエントリー酵素であるΔ12デサチュラーゼ遺伝子を有することを明らかにした。また、この遺伝子を相同組換えにより破壊した株は、Δ12デサチュラーゼの基質であるオレイン酸を野生株と比較して顕著に蓄積することや、産物であるリノール酸およびその生合成系の下流に位置するPUFAの減少が見られることを明らかにした。さらに、この株はエロンガーゼ/デサチュラーゼ経路によってPUFAを生産することが可能なため、PUFA-PKS経路遺伝子を破壊しても致死性にならないことも明らかにした。この報告は、ラビリンチュラ類でもPUFA生合成経路としてエロンガーゼ/デサチュラーゼ経路が機能していることを初めて示したもので、非特許文献6でも取り上げられた。
【0006】
冒頭で、主要な脂肪酸としてDHAを産生するラビリンチュラ類が産業的に広く利用されていることを述べた。これに対し、DHA以外の所望のPUFAを主要な脂肪酸として含有するラビリンチュラ類を作出することは、PUFA-PKS経路とエロンガーゼ/デサチュラーゼ経路の双方を有するラビリンチュラ類の発見およびこれらに対する形質転換技術(特許文献3、非特許文献7)の適用によって初めて可能になった。すなわち、まず相同組換えによる遺伝子破壊によりPUFA-PKS経路の遺伝子を破壊し、次いでエロンガーゼ/デサチュラーゼ経路を構成する酵素の遺伝子を適宜破壊または過剰発現することにより、DHA以外の特定のPUFAを主要な脂肪酸として含有する株を作出することが可能である。具体的には、特許文献3の実施例12が例示される。この実施例には、
Thraustochytrium
aureum
ATCC34304のPUFA-PKS経路の遺伝子を破壊し、次いでC20エロンガーゼ遺伝子を破壊し、さらにミズカビ
Saprolegnia
diclina
由来ω3デサチュラーゼ遺伝子を導入することにより、野生株と比較してアラキドン酸が約6倍、EPAが約10倍に増加し、かつDHAが約1/16倍に減少した株の作出に成功したことが記載されている。
【先行技術文献】
【特許文献】
【0007】
特許3127161号公報
特許3669372号公報
WO2012/043826号
米国特許公開US2005/0014231号公報
【非特許文献】
【0008】
ツヴィ・コーエン(Zvi Cohen)ら編,「シングル セル オイルズ マイクロバイアル アンド アルガル オイルズ セカンド エディション(Single Cel1 0ils Microbial and Algal Oils 2nd edition)」,(米国),エイオーシーエス・プレス(AOCS Press),2010年,p.88
Metz JG, Roessler P, Faccioti D, et al. Production of polyunsaturated fatty acids by polyketide synthases in both prokaryotes and eukaryotes. Science 2001;293:290-293
Ratledge C. Omega-3 biotechnology: Errors and omissions, Biotechnology Advances 30 (2012) 1746-1747
Lippmeier J.C. et al., Lipids, 44(7), 621-630(2009)
Matsuda T, Sakaguchi K, Hamaguchi R, Kobayashi T, Abe E, Hama Y, Hayashi M, Honda D, Okita Y, Sugimoto S, Okino N, Ito M. The analysis of delta12 fatty acid desaturase function revealed that two distinct pathways are active for the synthesis of polyunsaturated fatty acids in Thraustochytrium aureum ATCC34304. J. Lipid Res. 53(6):1210-1222 (2012)
ASBMB today June 2012 p.30
Sakaguchi K. et al., Versatile Transformation System That Is Applicable to both Multiple Transgene Expression and Gene Targeting for Thraustochytrids. Appl. Environ. Microbiol. 78(9):3193-3202 (2012)
Yazawa K., Lipids, 31, Supple. 297-300 (1996)
日本農芸化学会会誌, 77, 2, 150-153(2003)
「バイオ実験イラストレイテッド 第2巻 遺伝子解析の基礎」、p63-68、秀潤社、1995年発行
Sanger, F. et al., Proc. Natl. Acad. Sci, 74, 5463 (1977)
Cigan and Donahue, 1987; Romanos et al., 1992
Ausubel F.M. et al., Current Protocols in Molecular Biology, Unit 13(1994)」
Guthrie C., Fink G. et al., Methods in Enzymology: Guide to Yeast Genetics and Molecular Biology, Volume 194(1991)
Qiu, X., et al. J. Biol. Chem., 276, 31561-6 (2001)
Abe E., et al., J. Biochem, 140, 247-253 (2006)
「バイオ実験イラストレイテッド 第2巻 遺伝子解析の基礎」、p117-128、秀潤社、1995年発行
DIG説明書集[日本語版] 8th, Roche Applied Science
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、脂肪酸組成が改変するように遺伝子改変されたラビリンチュラ類、好ましくはエロンガーゼ/デサチュラーゼ経路のみでPUFAを生産するラビリンチュラ類、それらを用いたPUFAの製造方法およびそれらを用いて製造されたPUFAを含有する脂質(微生物油)およびそれらの使用を提供することを課題とする。
PUFA-PKS経路とエロンガーゼ/デサチュラーゼ経路の双方を有するラビリンチュラ類を用いて、DHA以外の特定のPUFAを主要な脂肪酸として含有する株を作出するためには、まずPUFA-PKS経路によるDHA生産を何らかの手段で停止あるいは抑制する必要があった。具体的には相同組換えによるPUFA-PKS経路の遺伝子の破壊の他に、UV照射や薬剤処理等による突然変異株の取得、RNAi(RNA interference)によるPUFA-PKS遺伝子の発現抑制等の方法が考えられる。
これらの方法のうち、相同組換えによる遺伝子破壊については先述のとおり、技術的には確立されている。しかし、マーカーとして使用可能な薬剤耐性遺伝子の数は一般に限りがあるので、PUFA-PKS経路の遺伝子の破壊にマーカーを用いると、さらなる遺伝子破壊や遺伝子導入の際に使用できるマーカーの数が減少するという問題があった。このことは、PUFA-PKS経路の遺伝子の破壊の後に実施可能な遺伝子破壊・遺伝子導入の回数の減少を意味するので、より高性能な株を作出する上での障害となる。
また、突然変異やRNAiによる発現抑制は他生物では実績がある方法だが、ラビリンチュラ類におけるこれらの方法による所望の株の取得可能性は不明である。
本発明は、内因性PUFA-PKS経路によるPUFAの生産能が無いあるいは極めて微弱であり、かつ内因性エロンガーゼ/デサチュラーゼ経路によるPUFA生産能を有するラビリンチュラ類、それらを用いたPUFAを含有する脂質の製造方法およびそれらを用いて製造したPUFAを含有する脂質を提供することを目的とするものである。
【課題を解決するための手段】
【0010】
本発明者らは鋭意検討の結果、内因性PUFA-PKS経路によるPUFAの生産能が無いあるいは極めて微弱であり、かつ内因性エロンガーゼ/デサチュラーゼ経路によるPUFA生産能を有するラビリンチュラ類が存在することを見出し、ラビリンチュラ類から得られる微生物油、微生物油産生ラビリンチュラ類、ならびに、それらの使用方法、および微生物油の使用に係る本発明を完成した。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
個人
酒類
1か月前
株式会社オシキリ
発酵検査装置
1か月前
個人
セルロース性物質の製造方法
9日前
松谷化学工業株式会社
澱粉分解物の製造方法
1か月前
株式会社GSP研究所
miRNA検出方法
20日前
東ソー株式会社
ポリマーコートビーズの製造方法
1か月前
インヒブルクス バイオサイエンシズ インコーポレイテッド
CLEC12a結合性ポリペプチド及びその使用
2か月前
株式会社写真化学
菌体観察装置
2か月前
サッポロビール株式会社
アルコール飲料
12日前
本田技研工業株式会社
培養システム
2か月前
サッポロビール株式会社
アルコール飲料
12日前
豊田合成株式会社
細胞培養膜及び細胞培養方法
2か月前
アサヒビール株式会社
容器詰麦芽発酵飲料
6日前
国立大学法人東京海洋大学
ウイルス不活化用組成物
2か月前
東ソー株式会社
アデノ随伴ウイルスのスクリーニング方法
1か月前
住友化学株式会社
積層体
2か月前
株式会社エムスタイル
微生物群の賦活化方法
20日前
学校法人君が淵学園
核酸の部位特異的アシル修飾剤
8日前
株式会社渡辺オイスター研究所
脂肪細胞の成長抑制剤
1か月前
三井化学株式会社
培養容器
1か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
5日前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
5日前
学校法人近畿大学
好気性微生物の培養方法および培養器
5日前
サムコ株式会社
ポアデバイス
9日前
日本バイリーン株式会社
細胞培養基材、および、その製造方法
3か月前
花王株式会社
抗アミラーゼ抗体
2か月前
住友化学株式会社
細胞チップ
2か月前
旭化成ファーマ株式会社
ジアホラーゼ
1か月前
東ソー株式会社
トリ骨髄芽細胞腫ウイルス逆転写酵素の製造方法
3か月前
国立大学法人富山大学
T細胞受容体およびその利用
9日前
株式会社豊田中央研究所
環境核酸モニタリング方法
5日前
三浦工業株式会社
試料液に含まれる微生物の生体の抽出方法
9日前
ウシオ電機株式会社
培養容器
1か月前
公立大学法人宮城大学
D-セリンの製造方法
1か月前
ZACROS株式会社
培養装置、及び培養装置積層体
1か月前
株式会社トクヤマ
乾燥用保護剤、生体試料測定試薬、及び分析方法
1か月前
続きを見る
他の特許を見る


 特許ウォッチ
特許ウォッチ