TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
公開番号
2024078959
公報種別
公開特許公報(A)
公開日
2024-06-11
出願番号
2022191608
出願日
2022-11-30
発明の名称
耐熱性タンパク質をスクリーニングするための方法およびキット
出願人
国立大学法人鳥取大学
代理人
個人
,
個人
,
個人
,
個人
主分類
C12N
15/09 20060101AFI20240604BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】標的タンパク質自体の酵素活性に依存する必要なく耐熱性タンパク質を同定できる、耐熱性タンパク質のスクリーニング技術を提供する。
【解決手段】複数の標的ポリペプチドをコードする複数の核酸を含む核酸ライブラリーで形質転換された好熱菌の集団を45℃以上のスクリーニング温度で培養する工程であって、複数の標的ポリペプチドは互いにアミノ酸配列の違いを有しており末端に50アミノ酸より短い共通配列の異種ペプチドを有する、工程と、異種ペプチドに結合するレポーターポリペプチドを上記集団に由来する複数のクローンの試料に添加する工程と、レポーターポリペプチドの結合量を検出して、結合量がより多い1つ以上のクローンを、スクリーニング温度に耐性である耐熱性標的ポリペプチドを蓄積させた好熱菌のクローンとして同定する工程とを含む、耐熱性ポリペプチドのスクリーニング方法、及びその方法を行うためのキット。
【選択図】なし
特許請求の範囲
【請求項1】
複数の標的ポリペプチドをコードする複数の核酸を含む核酸ライブラリーで形質転換された好熱菌の集団を、45℃以上のスクリーニング温度で培養する工程であって、ここで前記複数の標的ポリペプチドは、互いにアミノ酸配列の違いを有しており末端に50アミノ酸より短い共通配列の異種ペプチドを有する、工程と、
前記異種ペプチドに結合するレポーターポリペプチドを、前記好熱菌の集団に由来する複数のクローンの試料に添加する工程と、
前記複数のクローンの試料における前記レポーターポリペプチドの結合量を検出して、前記レポーターポリペプチドの結合量がより多い1つ以上のクローンを、前記スクリーニング温度に耐性である耐熱性標的ポリペプチドを蓄積させた好熱菌のクローンとして同定する工程と
を含む、耐熱性ポリペプチドのスクリーニング方法。
続きを表示(約 1,100 文字)
【請求項2】
前記好熱菌はGeobacillus属の好熱菌である、請求項1に記載のスクリーニング方法。
【請求項3】
前記レポーターポリペプチドの結合量は、前記レポーターポリペプチドが生じる酵素活性または蛍光の量として検出される、請求項1に記載のスクリーニング方法。
【請求項4】
前記異種ペプチドはスプリットレポーターの小断片であり、前記レポーターポリペプチドは前記スプリットレポーターの大断片である、請求項3に記載のスクリーニング方法。
【請求項5】
前記異種ペプチドはβ-ガラクトシダーゼのαフラグメントであり、前記レポーターポリペプチドは前記β-ガラクトシダーゼのωフラグメントであり、前記結合量は、前記αフラグメントと前記ωフラグメントのα相補性により生じる酵素活性の量として検出される、請求項4に記載のスクリーニング方法。
【請求項6】
前記異種ペプチドはスプリットGFPのGFP11フラグメントであって前記レポーターポリペプチドは前記スプリットGFPのGFP1-10フラグメントであるか、または、前記異種ペプチドは他のGFP由来スプリット蛍光タンパク質のGFP11フラグメントに対応するフラグメントであって前記レポーターポリペプチドは前記他のGFP由来スプリット蛍光タンパク質のGFP1-10フラグメントに対応するフラグメントであり、
前記結合量は、前記異種ペプチドと前記レポーターポリペプチドの自己会合により生じる蛍光の量として検出される、請求項4に記載のスクリーニング方法。
【請求項7】
前記培養する工程の前に、好熱菌の集団を前記核酸ライブラリーで形質転換する工程をさらに含む、請求項1に記載のスクリーニング方法。
【請求項8】
プラスミドベクターと、使用説明書とを含む、請求項1~7のいずれか一項に記載の方法を行うためのキットであって、
前記プラスミドベクターは、前記異種ペプチドをコードする核酸配列を含み、標的ポリペプチドをコードする核酸が挿入された場合に、前記異種ペプチドが末端に融合された標的ポリペプチドを前記好熱菌中で発現できるように構成され、
前記使用説明書は、前記方法の各工程を行うための指示を含む、キット。
【請求項9】
さらに、前記レポーターポリペプチドまたはそれをコードする核酸を含む、請求項8に記載のキット。
【請求項10】
さらに、前記形質転換される前の好熱菌、およびエラープローンDNAポリメラーゼのうちのいずれかまたは両方を含む、請求項8に記載のキット。
発明の詳細な説明
【技術分野】
【0001】
本開示は、遺伝子工学技術を利用したタンパク質の開発および生産の分野に関する。
続きを表示(約 4,200 文字)
【背景技術】
【0002】
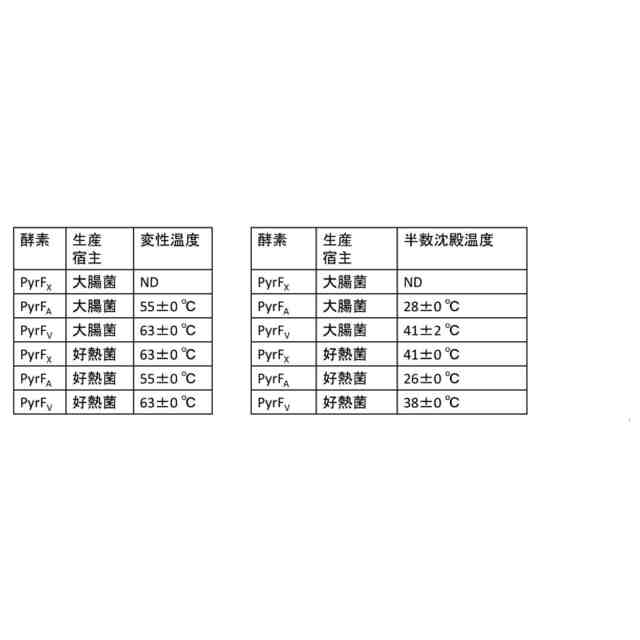
遺伝子工学技術により産生される人工タンパク質は産業的利用の大きな可能性を有しており、実際に医薬、触媒、材料等を含む様々な産業分野で既に広く利用されている。産業的に有用な人工タンパク質の開発、製造、および利用をしばしば妨げる要素の一つは、そのような人工タンパク質の不安定性、特に、熱に対する不安定性である。例えば、工業的な反応の触媒として利用するタンパク質(酵素)は、その反応が起こる温度(多くの場合、室温や生体温度より高い)において安定であることが求められるが、そのような温度で安定でありかつ必要な酵素活性を提供できるタンパク質は、天然にはそもそも稀であるし、天然タンパク質に基づいて人工的に開発することも容易ではない。どのアミノ酸残基を変異させれば耐熱性が向上するかという手掛かりは通常存在せず、圧倒的多数の変異は耐熱性の向上に繋がらないばかりかタンパク質の適正な折り畳みおよび機能にとって有害である。さらに、耐熱性が向上したごく少数の変異タンパク質をどのように同定するかということも難題である。例えば、ランダム変異させたライブラリーの中から1つずつクローンを分離してタンパク質を発現させ、その1つ1つのタンパク質の分析から耐熱性のものを見つけ出すというアプローチはきわめて非効率である。
【0003】
非特許文献1は、ランダム変異させたカナマイシンヌクレオチジルトランスフェラーゼ酵素遺伝子をBacillus stearothermophilus(現在はGeobacillus stearothermophilusに再分類されている)好熱菌に導入し、これらの好熱菌を高温かつカナマイシンの存在下で培養して、カナマイシン耐性クローンを選抜することにより、耐熱性が向上した変異酵素を同定したことを記載している。このスクリーニング方法は、耐熱性変異バリアントを同定するために、変異させたタンパク質自体の酵素活性に依存するものである。同様に、変異されたタンパク質自体の酵素活性に依存して、選択圧下の宿主細菌に生存能を付与できる耐熱性変異バリアントを同定した例として、非特許文献2がある。
【0004】
それに対して非特許文献3は、ランダム変異させた標的タンパク質を、非特許文献1の耐熱性カナマイシンヌクレオチジルトランスフェラーゼ酵素(レポーター)に融合させたものを、高温かつカナマイシンの存在下で培養された好熱菌Thermus thermophilusに発現させるスクリーニング実験を行い、該酵素レポーターの活性が維持されていたクローンを、耐熱性標的タンパク質を発現するクローンとして同定したことを記載している。このスクリーニング方法は、熱に不安定なため正しく折り畳まれない標的タンパク質は融合相手の酵素レポーターの高次折り畳みにも悪影響を及ぼすという原理に基づいている。この方法は、変異させたタンパク質自体の活性に依存せずに耐熱性変異バリアントを同定できるという利点を有する。しかし、250アミノ酸超という比較的大きなレポータータンパク質に融合させる必要性は制約となり得る。
【0005】
非特許文献4は、標的タンパク質を過剰発現させた後の大腸菌のコロニーを特殊なフィルター膜上に配置し、該大腸菌コロニーを高温に晒した後に、細胞を溶解させることを含む方法を記載している。この方法は、熱に不安定であるため折り畳みが壊れて凝集する標的タンパク質はフィルター膜上に残存する一方、上記高温でも可溶性を維持した標的タンパク質はフィルター膜を通過して、フィルター膜の下に配置されたニトロセルロース層上で捕捉され、検出されるという原理に基づいている。この方法はこの特殊なフィルター構造の使用に依存するものである。
【0006】
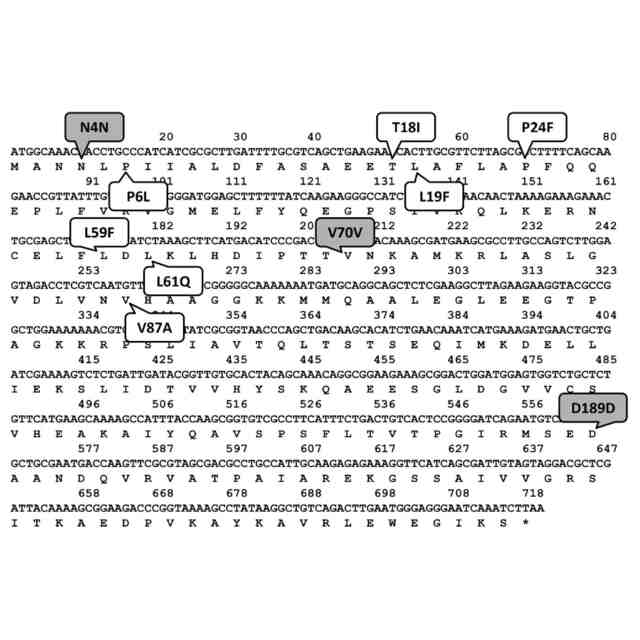
耐熱性タンパク質は熱だけでなく他の環境下でも頑強性を示す一般的傾向が見られ、耐熱性タンパク質を同定する方法の産業的有用性は高い。また耐熱性酵素は、例えば常温でもより高活性である、保存が容易である、使用寿命が長い等の利点を有し得る。また、耐熱性タンパク質の同定を通じて、例えば耐熱性を付与するアミノ酸残基の位置と種類を理解することにより、タンパク質の構造生物学的理解を深めることもできる。上記のように、改善された効率で耐熱性タンパク質をスクリーニングして同定するための巧妙な方法がいくつかは考案されてきたが、その数はまだ少数であるし、それらの方法もそれぞれ固有の制約を有している。いずれにせよ、耐熱性タンパク質をスクリーニングして同定することが難しい課題であるという一般的状況は継続している。
【先行技術文献】
【非特許文献】
【0007】
J. Biol. Chem. (1985) 260:15298-303
Appl. Environ. Microbiol. (2015) 81:149-58.
Nat. Methods (2007) 4:919-21
Nat. Commun. (2013) 4:2901
【発明の概要】
【発明が解決しようとする課題】
【0008】
本開示の実施形態は、従来技術の制約を少なくとも部分的に克服する、耐熱性タンパク質をスクリーニングするための新規技術を提供することを課題とする。特に、耐熱性タンパク質を同定するために標的タンパク質自体の酵素活性に依存する必要がない新規スクリーニング方法を提供することを課題とする。
【課題を解決するための手段】
【0009】
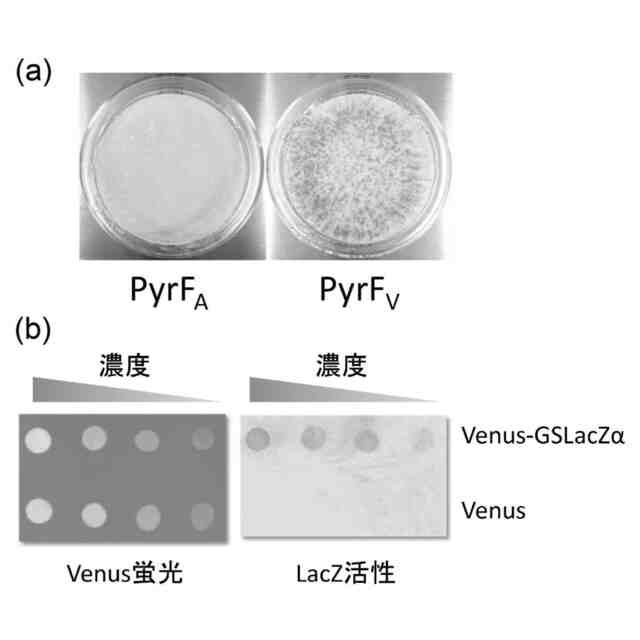
本発明者は、高温で生育する好熱菌で様々な異種タンパク質の発現を試みる研究のなかで、耐熱性のタンパク質は試料中に蓄積するのに対し、非耐熱性のタンパク質は、変性したりミスフォールディングした形態になるだけでなく、そういうミスフォールディングした形態のものがそもそも検出されず実質的に蓄積すらしないと見られる事象に気づいた。これは、きわめてシンプルでありながらこれまではっきりと認識されていなかった法則性である。本発明者は、この法則性を前提として耐熱性タンパク質のスクリーニングができるという仮説を立ててスクリーニング方法の開発を試みたところ、仮説に合致する観察と結果が得られ、本開示の発明をするに至った。
【0010】
本開示は以下の実施形態を含む。
[1]
複数の標的ポリペプチドをコードする複数の核酸を含む核酸ライブラリーで形質転換された好熱菌の集団を、45℃以上のスクリーニング温度で培養する工程であって、ここで前記複数の標的ポリペプチドは、互いにアミノ酸配列の違いを有しており末端に50アミノ酸より短い共通配列の異種ペプチドを有する、工程と、
前記異種ペプチドに結合するレポーターポリペプチドを、前記好熱菌の集団に由来する複数のクローンの試料に添加する工程と、
前記複数のクローンの試料における前記レポーターポリペプチドの結合量を検出して、前記レポーターポリペプチドの結合量がより多い1つ以上のクローンを、前記スクリーニング温度に耐性である耐熱性標的ポリペプチドを蓄積させた好熱菌のクローンとして同定する工程と
を含む、耐熱性ポリペプチドのスクリーニング方法。
[2]
前記好熱菌はGeobacillus属の好熱菌である、[1]に記載のスクリーニング方法。
[3]
前記レポーターポリペプチドの結合量は、前記レポーターポリペプチドが生じる酵素活性または蛍光の量として検出される、[1]または[2]に記載のスクリーニング方法。
[4]
前記異種ペプチドはスプリットレポーターの小断片であり、前記レポーターポリペプチドは前記スプリットレポーターの大断片である、[1]~[3]のいずれかに記載のスクリーニング方法。
[5]
前記異種ペプチドはβ-ガラクトシダーゼのαフラグメントであり、前記レポーターポリペプチドは前記β-ガラクトシダーゼのωフラグメントであり、前記結合量は、前記αフラグメントと前記ωフラグメントのα相補性により生じる酵素活性の量として検出される、[4]に記載のスクリーニング方法。
[6]
前記異種ペプチドはスプリットGFPのGFP11フラグメントであって前記レポーターポリペプチドは前記スプリットGFPのGFP1-10フラグメントであるか、または、前記異種ペプチドは他のGFP由来スプリット蛍光タンパク質のGFP11フラグメントに対応するフラグメントであって前記レポーターポリペプチドは前記他のGFP由来スプリット蛍光タンパク質のGFP1-10フラグメントに対応するフラグメントであり、
前記結合量は、前記異種ペプチドと前記レポーターポリペプチドの自己会合により生じる蛍光の量として検出される、[4]に記載のスクリーニング方法。
[7]
前記培養する工程の前に、好熱菌の集団を前記核酸ライブラリーで形質転換する工程をさらに含む、[1]~[6]のいずれかに記載のスクリーニング方法。
[8]
プラスミドベクターと、使用説明書とを含む、[1]~[7]のいずれか一項に記載の方法を行うためのキットであって、
前記プラスミドベクターは、前記異種ペプチドをコードする核酸配列を含み、標的ポリペプチドをコードする核酸が挿入された場合に、前記異種ペプチドが末端に融合された標的ポリペプチドを前記好熱菌中で発現できるように構成され、
前記使用説明書は、前記方法の各工程を行うための指示を含む、キット。
[9]
さらに、前記レポーターポリペプチドまたはそれをコードする核酸を含む、[8]に記載のキット。
[10]
さらに、前記形質転換される前の好熱菌、およびエラープローンDNAポリメラーゼのうちのいずれかまたは両方を含む、[8]または[9]に記載のキット。
【図面の簡単な説明】
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
国立大学法人鳥取大学
熱電変換素子及びその製造方法並びに熱電変換デバイス
3か月前
国立大学法人鳥取大学
過回転抑制機構を備えた垂直軸風車発電ユニット、及び過回転抑制機構を備えた垂直軸風車発電ユニットの制御方法
25日前
個人
酒類
1か月前
株式会社オシキリ
発酵検査装置
25日前
松谷化学工業株式会社
澱粉分解物の製造方法
23日前
東ソー株式会社
ポリマーコートビーズの製造方法
24日前
株式会社GSP研究所
miRNA検出方法
2日前
インヒブルクス バイオサイエンシズ インコーポレイテッド
CLEC12a結合性ポリペプチド及びその使用
1か月前
株式会社写真化学
菌体観察装置
2か月前
株式会社雷神の風
試料採取装置
2か月前
本田技研工業株式会社
培養システム
1か月前
豊田合成株式会社
細胞培養膜及び細胞培養方法
1か月前
住友化学株式会社
積層体
1か月前
株式会社エムスタイル
微生物群の賦活化方法
2日前
東ソー株式会社
アデノ随伴ウイルスのスクリーニング方法
1か月前
国立大学法人東京海洋大学
ウイルス不活化用組成物
1か月前
三井化学株式会社
培養容器
1か月前
株式会社渡辺オイスター研究所
脂肪細胞の成長抑制剤
24日前
住友化学株式会社
細胞チップ
1か月前
日本バイリーン株式会社
細胞培養基材、および、その製造方法
2か月前
花王株式会社
抗アミラーゼ抗体
2か月前
旭化成ファーマ株式会社
ジアホラーゼ
1か月前
東ソー株式会社
トリ骨髄芽細胞腫ウイルス逆転写酵素の製造方法
2か月前
国立大学法人 宮崎大学
水素発生装置
1か月前
ZACROS株式会社
培養装置、及び培養装置積層体
1か月前
株式会社トクヤマ
乾燥用保護剤、生体試料測定試薬、及び分析方法
25日前
ウシオ電機株式会社
培養容器
1か月前
公立大学法人宮城大学
D-セリンの製造方法
23日前
雪印メグミルク株式会社
D-セリン濃度低減用組成物
23日前
セイコーエプソン株式会社
糖化酵素の回収方法
1か月前
グリーンケミカルズ株式会社
フェノール組成物およびポリマー
18日前
ポーラ化成工業株式会社
Piezo遺伝子発現量亢進剤
2か月前
花王株式会社
歯周病リスクの検出方法
2か月前
サントリーホールディングス株式会社
ビールテイスト飲料
1か月前
学校法人近畿大学
新規酵母と柿を用いた酒類の製造方法
1か月前
株式会社システマックス
植物細胞外小胞、その調製方法及び用途
2か月前
続きを見る
他の特許を見る
 特許ウォッチ
特許ウォッチ