TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2024037852
公報種別
公開特許公報(A)
公開日
2024-03-19
出願番号
2023209381,2023094901
出願日
2023-12-12,2017-12-20
発明の名称
融合ポリペプチド、医薬組成物、核酸、宿主細胞、及び融合ポリペプチドを作成する方法
出願人
キュー バイオファーマ, インコーポレイテッド
代理人
個人
,
個人
主分類
C12N
15/62 20060101AFI20240312BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】変異免疫調節ポリペプチド、及び変異免疫調節ペプチドを含む融合ポリペプチドを提供する。
【解決手段】a)複数の変異IL-2ポリペプチドと、b)異種融合パートナーとを含んでおり、複数の変異IL-2ポリペプチドは、第1の変異IL-2ポリペプチド及び第2の変異IL-2ポリペプチドを含み、第1の変異IL-2ポリペプチド及び第2の変異IL-2ポリペプチドは、特定のアミノ酸配列を有するアミノ酸配列を含み、アミノ酸16はAlaであって、アミノ酸42はAlaであり、第1の変異IL-2ポリペプチド及び第2の変異IL-2ポリペプチドは、リンカーにより連結されており、異種融合パートナーは、主要組織適合抗原複合体(MHC)ポリペプチドを含まず、異種融合パートナーは、抗体Fc領域である、融合ポリペプチドを提供する。
【選択図】図1
特許請求の範囲
【請求項1】
融合ポリペプチドであって、
a)複数の変異IL-2ポリペプチドと、
b)異種融合パートナーと
を含んでおり、
前記複数の変異IL-2ポリペプチドは、第1の変異IL-2ポリペプチド及び第2の変異IL-2ポリペプチドを含み、
前記第1の変異IL-2ポリペプチド及び第2の変異IL-2ポリペプチドは、配列番号44に示されるアミノ酸配列を有するアミノ酸配列を含み、
アミノ酸16はAlaであって、アミノ酸42はAlaであり、
前記第1の変異IL-2ポリペプチド及び第2の変異IL-2ポリペプチドは、リンカーにより連結されており、
前記異種融合パートナーは、主要組織適合抗原複合体(MHC)ポリペプチドを含まず、
前記異種融合パートナーは、抗体Fc領域である、
融合ポリペプチド。
続きを表示(約 560 文字)
【請求項2】
請求項1に記載の融合ポリペプチドを含む、医薬組成物。
【請求項3】
請求項1に記載の融合ポリペプチドをコードするヌクレオチド配列を含む、核酸。
【請求項4】
請求項3に記載の核酸を含む、宿主細胞。
【請求項5】
融合ポリペプチドを作成する方法であって、
請求項4に記載の宿主細胞を、前記宿主細胞が前記融合ポリペプチドを産生するように、培養することを含む、
方法。
【請求項6】
前記複数の変異IL-2ポリペプチドは、第1の変異IL-2ポリペプチド及び第2の変異IL-2ポリペプチドからなる、請求項1に記載の融合ポリペプチド。
【請求項7】
請求項6に記載の融合ポリペプチドを含む、医薬組成物。
【請求項8】
請求項6に記載の融合ポリペプチドをコードするヌクレオチド配列を含む、核酸。
【請求項9】
請求項8に記載の核酸を含む、宿主細胞。
【請求項10】
融合ポリペプチドを作成する方法であって、
請求項9に記載の宿主細胞を、前記宿主細胞が前記融合ポリペプチドを産生するように、培養することを含む、
方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【背景技術】
【0001】
適応免疫応答は、T細胞の表面に存在するT細胞受容体(TCR)と、主要組織適合抗
原複合体(MHC;ヒトにおいてはヒト白血球抗原(HLA)複合体とも称される)によ
って抗原提示細胞(APC)の表面に非共有結合で存在する小さなペプチド抗原との結合
を伴う。この結合は、免疫系の標的化機構に相当するものであり、T細胞の調節(活性化
または抑制)及びエフェクター機能に必要な分子相互作用である。エピトープ特異的な細
胞標的化に続き、APCに見られる共刺激タンパク質と、T細胞の対応する共刺激タンパ
ク質との結合を介して標的化T細胞が活性化される。
続きを表示(約 7,400 文字)
【0002】
T細胞の特異性及び活性化または抑制を誘導するためには、エピトープ/TCR結合と
、APCの共刺激タンパク質及びT細胞の共刺激タンパク質の結合との両方のシグナルが
必要である。TCRは所与のエピトープに特異的であるが、共刺激タンパク質はエピトー
プ特異的ではなく、代わりに、すべてのT細胞または大きなT細胞サブセットに一般に発
現する。
【発明の概要】
【0003】
本開示は、変異免疫調節ポリペプチド、及び変異免疫調節ペプチドを含む融合ポリペプ
チドを提供する。本開示は、T細胞調節多量体ポリペプチド、及びT細胞調節多量体ポリ
ペプチドを含む組成物を提供し、T細胞調節多量体ポリペプチドは、本開示の変異免疫調
節ポリペプチドを含む。
【0004】
本開示は、T細胞調節多量体ポリペプチドをコードするヌクレオチド配列を含む核酸、
及び核酸を含む宿主細胞を提供する。本開示は、T細胞の活性の調節方法を提供し、この
方法は、T細胞と、本開示のT細胞調節多量体ポリペプチドとの接触を含む。
【図面の簡単な説明】
【0005】
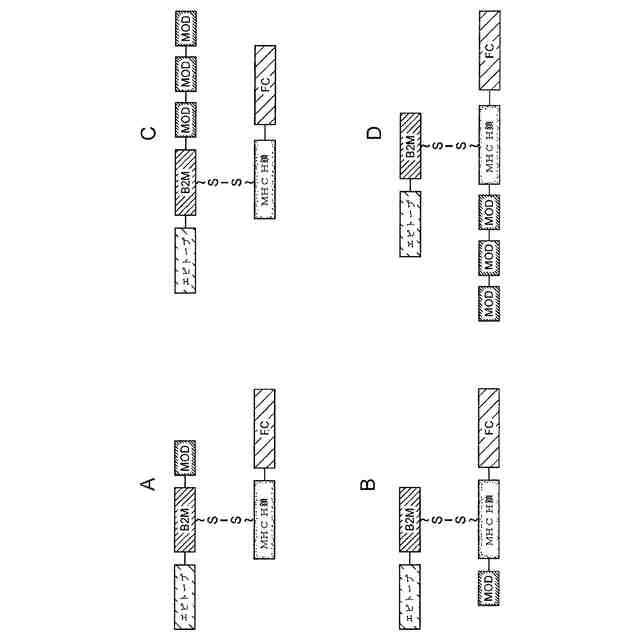
A~Dは、本開示のT細胞調節多量体ポリペプチドのさまざまな実施形態を模式的に示す。これらの実施形態では、別々のポリペプチドに存在するMHC(例えば、HLA)ポリペプチドの間にジスルフィド結合が形成される。
A~Dは、野生型ヒトIL-2のアミノ酸配列(A)、及び変異IL-2ポリペプチドのアミノ酸配列(B~D)を示す。
E~Hは、変異IL-2ポリペプチドのアミノ酸配列を示す。
I~Lは、変異IL-2ポリペプチドのアミノ酸配列を示す。
M~Pは、変異IL-2ポリペプチドのアミノ酸配列を示す。
Qは、変異IL-2ポリペプチドのアミノ酸配列を示す。
A及びBは、IL-2受容体のアルファ鎖、及びIL-2受容体のベータ鎖のアミノ酸配列を示す。
Cは、IL-2受容体のガンマ鎖のアミノ酸配列を示す。
免疫グロブリンFcポリペプチドのアミノ酸配列を示す。
免疫グロブリンFcポリペプチドのアミノ酸配列を示す。
免疫グロブリンFcポリペプチドのアミノ酸配列を示す。
A及びBは、ヒト白血球抗原(HLA)クラスI重鎖ポリペプチドのアミノ酸配列を示す。シグナル配列には下線が引かれている。
Cは、ヒト白血球抗原(HLA)クラスI重鎖ポリペプチドのアミノ酸配列を示す。シグナル配列には下線が引かれている。
Homo sapiens(NP_004039.1;配列番号95)、Pan troglodytes(NP_001009066.1;配列番号96)、Macaca mulatta(NP_001040602.1;配列番号97)、Bos Taurus(NP_776318.1;配列番号98)、及びMus musculus(NP_033865.2;配列番号99)に由来するベータ-2ミクログロブリン(β2M)前駆体(すなわち、リーダー配列を含む)の複数のアミノ酸配列のアライメントを示す。アミノ酸1~20は、シグナルペプチドである。
一過性の遺伝子導入の後の本開示のIL-2/synTac(「Cue-IL-2-a」及び「Cue-IL-2-b」)の産生(未精製収量)を示す。
一過性の遺伝子導入の後の本開示のIL-2/synTac(「Cue-IL-2-a」及び「Cue-IL-2-b」)の産生(精製産物)を示す。
本開示のIL-2/synTacの産生を示し、これらのIL-2/synTacでは、IL-2ポリペプチドは、軽鎖(MHCクラスI分子の軽鎖(例えば、β2M)を含むポリペプチド鎖)または重鎖(MHCクラスI分子の重鎖を含むポリペプチド鎖)に存在する。
本開示のIL-2/synTacの産生を示し、これらのIL-2/synTacでは、IL-2ポリペプチドは、軽鎖(MHCクラスI分子の軽鎖(例えば、β2M)を含むポリペプチド鎖)または重鎖(MHCクラスI分子の重鎖を含むポリペプチド鎖)に存在する。
IL-2/syn-Tacの発現レベルを示し、これらのIL-2/syn-Tacでは、IL-2は、野生型(wt)であるか、またはF42A、D20K、Q126A、E15A、Y45A、及びH16Aのさまざまな組み合わせを含む。
本開示のIL-2/synTacの発現を示し、これらのIL-2/synTacでは、IL-2の1つのコピー(1X)、2つのコピー(2X)、または3つのコピー(3X)がsynTacに存在する。
本開示のIL-2/synTacによる抗原特異的CD8
+
T細胞及び非特異的CD8
+
T細胞のインビトロの刺激を示し、ここでは、F42A置換及びH16A置換を含むIL-2変異体の2つのコピーがsynTacに存在する。
特異的(リンパ球性脈絡髄膜炎ウイルス(LCMV))CD8
+
T細胞または非特異的(OT1(オボアルブミン認識))CD8
+
T細胞に対するIL-2/synTacの結合を示す。
抗原特異的(LCMV)CD8
+
T細胞または非特異的(BL6)CD8
+
T細胞におけるIL-2/synTac介在性のシグナル伝達を示す。
本開示のIL-2/synTacをさまざまなIL-2/synTac濃度で用いてCD8
+
抗原特異的(LCMV)細胞またはCD8
+
非特異的(BL6)細胞を刺激した後のリン酸化シグナル伝達兼転写活性化因子5(pSTAT5)陽性細胞のパーセントを示す。
本開示のIL-2/synTacをさまざまなIL-2/synTac濃度で用いてCD8
+
抗原特異的(LCMV)細胞またはCD8
+
非特異的(BL6)細胞を刺激した後のリン酸化シグナル伝達兼転写活性化因子5(pSTAT5)陽性細胞のパーセントを示す。
本開示のIL-2/synTacをさまざまなIL-2/synTac濃度で用いてCD8
+
抗原特異的(LCMV)細胞またはCD8
+
非特異的(BL6)細胞を刺激した後のリン酸化シグナル伝達兼転写活性化因子5(pSTAT5)陽性細胞のパーセントを示す。
本開示のIL-2/synTacをさまざまなIL-2/synTac濃度で用いてCD8
+
抗原特異的(LCMV)細胞またはCD8
+
非特異的(BL6)細胞を刺激した後のリン酸化シグナル伝達兼転写活性化因子5(pSTAT5)陽性細胞のパーセントを示す。
本開示のIL-2/synTacをさまざまなIL-2/synTac濃度で用いてCD8
+
抗原特異的(LCMV)細胞またはCD8
+
非特異的(BL6)細胞を刺激した後のリン酸化シグナル伝達兼転写活性化因子5(pSTAT5)陽性細胞のパーセントを示す。
本開示のIL-2/synTacをさまざまなIL-2/synTac濃度で用いてCD8
+
抗原特異的(LCMV)細胞またはCD8
+
非特異的(BL6)細胞を刺激した後のリン酸化シグナル伝達兼転写活性化因子5(pSTAT5)陽性細胞のパーセントを示す。
本開示のIL-2/synTacのインビボの活性を示す。左パネルは、リン酸緩衝生理食塩水(PBS)、組換えIL-2(rIL-2)、または本開示のIL-2/synTacを投与した後の抗原特異的CD8
+
T細胞の数の倍率変化を示す。右パネルは、PBS、rIL-2、または本開示のIL-2/synTacを投与した後の抗原特異的な応答及び抗原非特異的な応答を示す。
用量漸増による影響を示す。
投与経路による影響を示す。
腫瘍に対するインビボの効力に与えるIL-2のコピー数の影響を示す。
腫瘍に対するインビボの効力に与えるIL-2のコピー数の影響を示す。
本開示のIL-2/synTacを10mg/kgのIL-2/synTac量で腹腔内投与した後の、その血清半減期を示す。
本開示のIL-2/synTacを10mg/kgのIL-2/synTac量で腹腔内投与してから2時間後の、その安定性を示す。
本開示のIL-2/synTacを4℃または37℃で5日間保持した後のIL-2/synTacに関するサイズ排除クロマトグラフィーのデータを示す。
リーダーペプチドを含む本開示のIL-2/synTacの重鎖のアミノ酸配列を示し、IL-2/synTac重鎖は、N297A置換を有するIgG1 Fcを含む。
リーダーペプチドを含まない本開示のIL-2/synTacの重鎖のアミノ酸配列を示し、IL-2/synTac重鎖は、N297A置換を有するIgG1 Fcを含む。
図21に示されるIL-2/synTac重鎖をコードするヌクレオチド配列を示す。
図23Aの配列に対する説明を示す。
リーダーペプチドを含む本開示のIL-2/synTacの重鎖のアミノ酸配列を示し、IL-2/synTac重鎖は、L234A置換及びL235A置換を有するIgG1 Fcを含む。
リーダーペプチドを含まない本開示のIL-2/synTacの重鎖のアミノ酸配列を示し、IL-2/synTac重鎖は、L234A置換及びL235A置換を有するIgG1 Fcを含む。
図24に示されるIL-2/synTac重鎖をコードするヌクレオチド配列を示す。
図26Aの配列に対する説明を示す。
リーダーペプチドを含む本開示のIL-2/synTacの重鎖のアミノ酸配列を示し、IL-2/synTac重鎖は、L234F置換、L235E置換、及びP331S置換を有するIgG1 Fcを含む。
リーダーペプチドを含まない本開示のIL-2/synTacの重鎖のアミノ酸配列を示し、IL-2/synTac重鎖は、L234F置換、L235E置換、及びP331S置換を有するIgG1 Fcを含む。
図27に示されるIL-2/synTac重鎖をコードするヌクレオチド配列を示す。
図29Aの配列に対する説明を示す。
リーダーペプチドを含む本開示のIL-2/synTacの軽鎖のアミノ酸配列を示し、IL-2/synTac軽鎖は、ヒトパピローマウイルス(HPV)E7エピトープを含む。
リーダーペプチドを含まない本開示のIL-2/synTacの軽鎖のアミノ酸配列を示し、IL-2/synTac軽鎖は、HPV E7エピトープを含む。
図30に示されるIL-2/synTac軽鎖をコードするヌクレオチド配列を示す。
A~Dは、野生型ヒトIgG1 Fcのアミノ酸配列(A)、L234F置換、L235E置換、及びP331S置換を有するIgG1 Fcのアミノ酸配列(B)、N297A置換を有するIgG1 Fcのアミノ酸配列(C)、ならびにL234A置換及びL235A置換を有するIgG1 Fcのアミノ酸配列(D)を示す。
A~Cは、β2-ミクログロブリン(R12C)ポリペプチドのアミノ酸配列(A)、変異IL-2(H16A、F42A)ポリペプチドのアミノ酸配列(B)、及びクラスI MHC-H鎖A0201(Y84A、A236C)のアミノ酸配列(C)を示す。
ヒトCMV特異的CD8
+
T細胞のIL-2/synTac介在性の増殖を示す。
変異IL-2ポリペプチドを含むsynTacの発現データ及び受容体結合データを示す。
HPV16 E7(11~20)特異的初代ヒトCD8
+
T細胞に対するIL-2/synTacの結合をフローサイトメトリーによって検出したものを示す。
HPV16 E7(11~20)特異的初代ヒトCD8
+
T細胞に対する変異IL-2/synTacの結合がSLP76のリン酸化に与える影響を示す。
HPV16 E7(11~20)特異的初代ヒトT細胞に対する変異IL-2/synTacの結合がCD25、グランザイムB、及びCD107αの産生に与える影響を示す。
HPV16 E7(11~20)特異的初代ヒトCD8
+
T細胞に対する変異IL-2/synTacの結合がIFN-γの産生に与える影響を示す。
【発明を実施するための形態】
【0006】
「ポリヌクレオチド」及び「核酸」という用語は、本明細書で互換的に使用され、リボ
ヌクレオチドまたはデオキシリボヌクレオチドのいずれかであり、長さが任意の重合形態
のヌクレオチドを指す。したがって、この用語は、限定はされないが、一本鎖、二本鎖、
または多鎖のDNAまたはRNA、ゲノムDNA、cDNA、DNAとRNAとのハイブ
リッド、あるいはプリン塩基及びピリミジン塩基を含むポリマー、あるいは他の天然のヌ
クレオチド塩基、化学的もしくは生化学的に改変されたヌクレオチド塩基、非天然のヌク
レオチド塩基、または誘導体化されたヌクレオチド塩基を含むポリマーを含む。
【0007】
「ペプチド」、「ポリペプチド」、及び「タンパク質」という用語は、本明細書で互換
的に使用され、化学的または生化学的に改変または誘導体化されたアミノ酸であるコード
アミノ酸及び非コードアミノ酸を含み得、長さが任意の重合形態のアミノ酸と、改変され
たペプチド骨格を有するポリペプチドとを指す。
【0008】
ポリヌクレオチドまたはポリペプチドは、別のポリヌクレオチドまたはポリペプチドに
対してある一定パーセントの「配列同一性」を有し、このことは、アライメントがとられ
、2つの配列が比較されると、その割合の塩基またはアミノ酸が同一であり、同一の相対
位置に存在することを意味する。配列同一性は、多くの異なる方法において決定すること
ができる。配列同一性を決定するために、さまざまな簡便な方法及びコンピュータープロ
グラム(例えば、BLAST、T-COFFEE、MUSCLE、MAFFTなど)を使
用して配列のアライメントをとることができ、こうした方法及びコンピュータープログラ
ムは、ncbi.nlm.nili.gov/BLAST、ebi.ac.uk/Too
ls/msa/tcoffee/、ebi.ac.uk/Tools/msa/musc
le/、mafft.cbrc.jp/alignment/software/を含む
、いくつかのサイトでワールドワイドウェブを通じて利用可能である。例えば、Alts
chul et al.(1990),J.Mol.Bioi.215:403-10を
参照のこと。
【0009】
「保存的アミノ酸置換」という用語は、同様の側鎖を有するアミノ酸残基の、タンパク
質における互換性を指す。例えば、脂肪族側鎖を有するアミノ酸の一群は、グリシン、ア
ラニン、バリン、ロイシン、及びイソロイシンからなり、脂肪族-ヒドロキシル側鎖を有
するアミノ酸の一群は、セリン及びスレオニンからなり、アミド含有側鎖を有するアミノ
酸の一群は、アスパラギン及びグルタミンからなり、芳香族側鎖を有するアミノ酸の一群
は、フェニルアラニン、チロシン、及びトリプトファンからなり、塩基性側鎖を有するア
ミノ酸の一群は、リジン、アルギニン、及びヒスチジンからなり、酸性側鎖を有するアミ
ノ酸の一群は、グルタミン酸及びアスパラギン酸からなり、含硫側鎖を有するアミノ酸の
一群は、システイン及びメチオニンからなる。保存的なアミノ酸置換群の例は、バリン-
ロイシン-イソロイシン、フェニルアラニン-チロシン、リジン-アルギニン、アラニン
-バリン-グリシン、及びアスパラギン-グルタミンである。
【0010】
(例えば、T細胞に存在するポリペプチド(例えば、T細胞受容体)に対する本開示の
T細胞調節多量体ポリペプチドの結合に関して)本明細書で使用される「結合」は、介在
する非共有結合的な相互作用を指す。結合相互作用は、解離定数(K
D
)が10
-6
M未満
、10
-7
M未満、10
-8
M未満、10
-9
M未満、10
-10
M未満、10
-11
M未満、10
-12
M未満、10
-13
M未満、10
-14
M未満、または10
-15
M未満であることによっ
て一般に特徴付けられる。「親和性」は、結合強度を指し、結合親和性の上昇は、K
D
の
低下と相関する。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
個人
酒類
1か月前
株式会社オシキリ
発酵検査装置
1か月前
個人
セルロース性物質の製造方法
7日前
松谷化学工業株式会社
澱粉分解物の製造方法
1か月前
東ソー株式会社
ポリマーコートビーズの製造方法
1か月前
株式会社GSP研究所
miRNA検出方法
18日前
株式会社写真化学
菌体観察装置
2か月前
インヒブルクス バイオサイエンシズ インコーポレイテッド
CLEC12a結合性ポリペプチド及びその使用
2か月前
本田技研工業株式会社
培養システム
2か月前
豊田合成株式会社
細胞培養膜及び細胞培養方法
1か月前
サッポロビール株式会社
アルコール飲料
10日前
サッポロビール株式会社
アルコール飲料
10日前
アサヒビール株式会社
容器詰麦芽発酵飲料
4日前
株式会社エムスタイル
微生物群の賦活化方法
18日前
東ソー株式会社
アデノ随伴ウイルスのスクリーニング方法
1か月前
学校法人君が淵学園
核酸の部位特異的アシル修飾剤
6日前
国立大学法人東京海洋大学
ウイルス不活化用組成物
2か月前
住友化学株式会社
積層体
1か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
3日前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
3日前
三井化学株式会社
培養容器
1か月前
株式会社渡辺オイスター研究所
脂肪細胞の成長抑制剤
1か月前
サムコ株式会社
ポアデバイス
7日前
花王株式会社
抗アミラーゼ抗体
2か月前
住友化学株式会社
細胞チップ
1か月前
学校法人近畿大学
好気性微生物の培養方法および培養器
3日前
日本バイリーン株式会社
細胞培養基材、および、その製造方法
2か月前
株式会社豊田中央研究所
環境核酸モニタリング方法
3日前
旭化成ファーマ株式会社
ジアホラーゼ
1か月前
国立大学法人富山大学
T細胞受容体およびその利用
7日前
ZACROS株式会社
培養装置、及び培養装置積層体
1か月前
三浦工業株式会社
試料液に含まれる微生物の生体の抽出方法
7日前
株式会社トクヤマ
乾燥用保護剤、生体試料測定試薬、及び分析方法
1か月前
国立大学法人 宮崎大学
水素発生装置
2か月前
セイコーエプソン株式会社
糖化酵素の回収方法
2か月前
雪印メグミルク株式会社
D-セリン濃度低減用組成物
1か月前
続きを見る
他の特許を見る


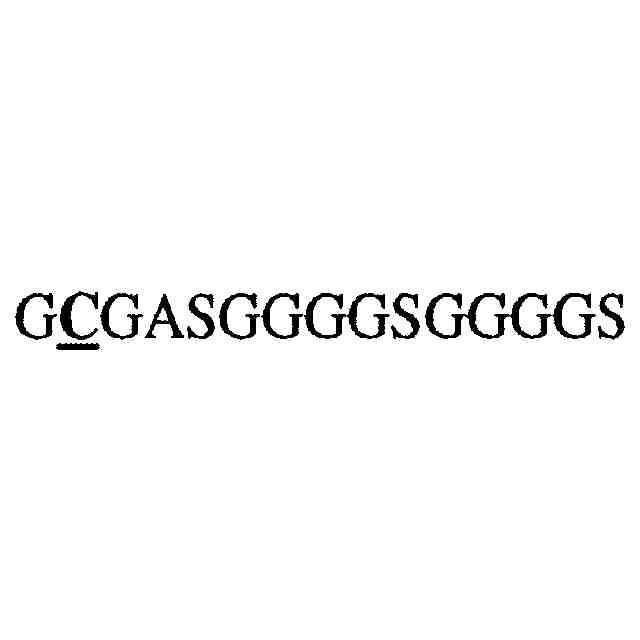







 特許ウォッチ
特許ウォッチ