TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2024156665
公報種別
公開特許公報(A)
公開日
2024-11-06
出願番号
2024106599,2021535281
出願日
2024-07-02,2019-12-18
発明の名称
タンパク質をコードするRNA
出願人
ヴェルサメブ アーゲー
代理人
園田・小林弁理士法人
主分類
C12N
15/62 20060101AFI20241029BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】タンパク質およびシグナルペプチドをコードする核酸配列を含むmRNA、ならびにタンパク質およびシグナルペプチドをコードする核酸を含む転写単位、発現ベクターまたは遺伝子治療用ベクターを提供する。
【解決手段】5’から3’方向に以下の順番で、i)シグナルペプチドおよびii)タンパク質をコードする核酸配列を含むmRNAであって、シグナルペプチドのアミノ酸配列のN末端のアミノ酸1~9は、2.2に等しいかまたはそれを超える平均疎水性スコアを有し、疎水性スコアは、カイト-デューリトルのスケールにより計算され、かつ、シグナルペプチドは、前記タンパク質にとって異種のシグナルペプチドであり、シグナルペプチドは、長さ16から40アミノ酸のアミノ酸配列からなり、ただし前記タンパク質はオキシドレダクターゼではない、mRNAが提供される。
【選択図】なし
特許請求の範囲
【請求項1】
タンパク質およびシグナルペプチドをコードする核酸配列を含むmRNAであって、
シグナルペプチドのアミノ酸配列のN末端のアミノ酸1~9は、2を超える平均疎水性スコアを有し、シグナルペプチドは、
i)前記タンパク質にとって異種のシグナルペプチドであって、前記タンパク質にとって異種のシグナルペプチドは、必要に応じて、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されており、ただし前記タンパク質はオキシドレダクターゼではない、前記タンパク質にとって異種のシグナルペプチド;
ii)前記タンパク質に相同なシグナルペプチドであって、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、前記タンパク質に相同なシグナルペプチド;および
iii)自然状態でシグナルペプチドの機能を有さない天然に存在するアミノ酸配列であって、必要に応じて少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、天然に存在するアミノ酸配列
からなる群から選択される、mRNA。
続きを表示(約 1,000 文字)
【請求項2】
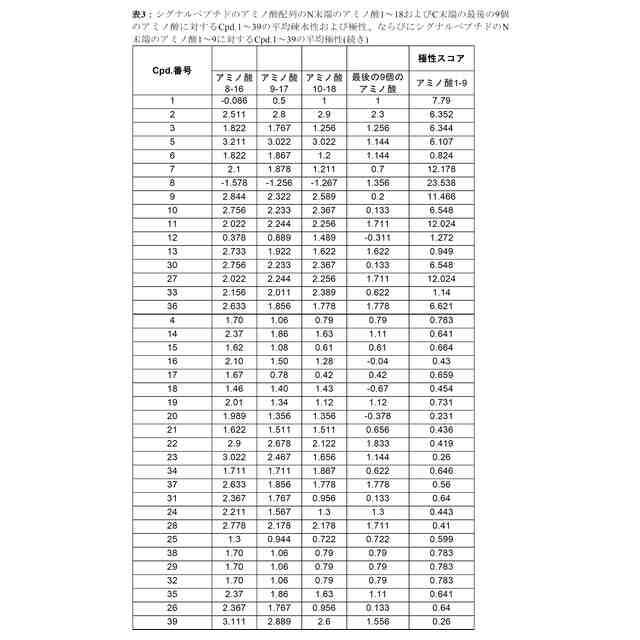
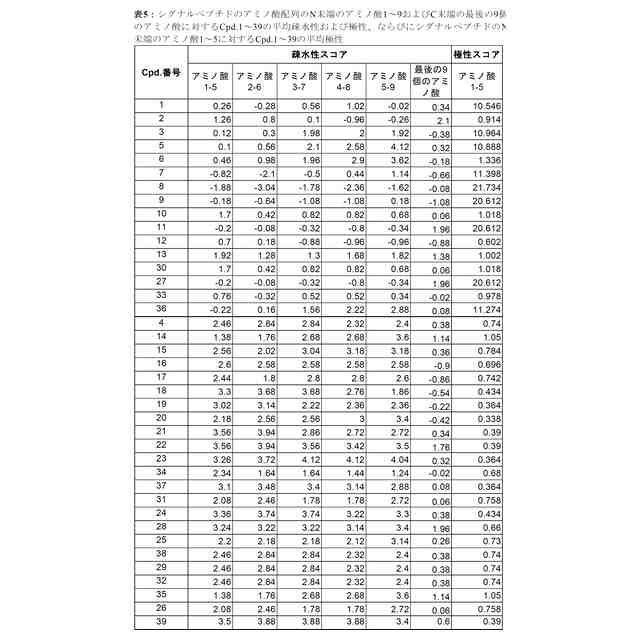
シグナルペプチドのアミノ酸配列のN末端のアミノ酸1~9が、6.1またはそれ未満の平均極性を有する、請求項1に記載のmRNA。
【請求項3】
シグナルペプチドのアミノ酸配列のC末端の最後の9つのアミノ酸の平均疎水性スコアが、シグナルペプチドのアミノ酸配列のN末端のアミノ酸1~9の平均疎水性スコアより少なくとも1.0単位低い、請求項1または2に記載のmRNA。
【請求項4】
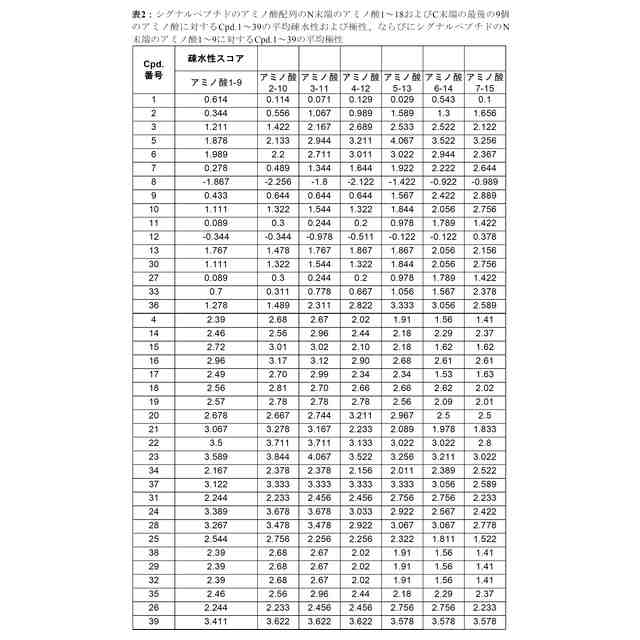
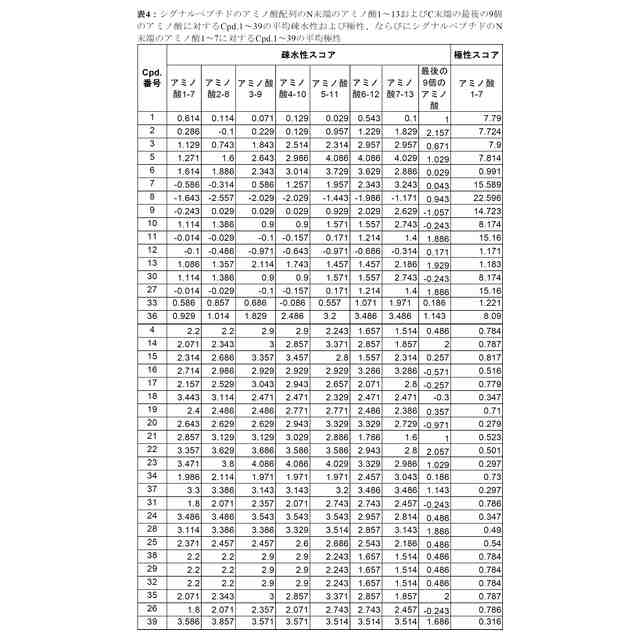
シグナルペプチドのアミノ酸配列のN末端のアミノ酸1~9、アミノ酸2~10、アミノ酸3~11、アミノ酸4~12およびアミノ酸5~13が、それぞれ1.5を超える平均疎水性スコアを有する、請求項1から3のいずれか一項に記載のmRNA。
【請求項5】
シグナルペプチドのアミノ酸配列のN末端のアミノ酸8~16の平均疎水性スコアが、シグナルペプチドのアミノ酸配列のN末端のアミノ酸3~11の平均疎水性スコアに等しいかまたはそれより低い、請求項1から4のいずれか一項に記載のmRNA。
【請求項6】
シグナルペプチドが、長さ18~40アミノ酸のアミノ酸配列を含み、シグナルペプチドのアミノ酸配列のN末端のアミノ酸10~18の平均疎水性スコアは、シグナルペプチドのアミノ酸配列のN末端のアミノ酸3~11の平均疎水性スコアより少なくとも0.5単位低い、請求項1から4のいずれか一項に記載のmRNA。
【請求項7】
シグナルペプチドのアミノ酸配列のC末端の最後の9つのアミノ酸の平均疎水性スコアが、シグナルペプチドのアミノ酸配列のN末端のアミノ酸3~11の平均疎水性スコアより少なくとも1.5単位低い、請求項1から6のいずれか一項に記載のmRNA。
【請求項8】
シグナルペプチドのアミノ酸配列の任意の9つの連続するアミノ酸の平均疎水性スコアが、4.1を超えない、請求項1から7のいずれか一項に記載のmRNA。
【請求項9】
シグナルペプチドのアミノ酸配列のC末端の最後の9つのアミノ酸が、負の疎水性スコアを有する少なくとも1つのアミノ酸を含む、請求項1から8のいずれか一項に記載のmRNA。
【請求項10】
負の疎水性スコアを有する少なくとも1つのアミノ酸が、G、Q、N、T、S、R、K、H、D、E、P、YおよびWからなる群から選択される、請求項9に記載のmRNA。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、タンパク質およびシグナルペプチドをコードする核酸配列を含むmRNAであって、シグナルペプチドのアミノ酸配列のN末端のアミノ酸1~9は、2を超える平均疎水性スコアを有し、シグナルペプチドは、
i)前記タンパク質にとって異種のシグナルペプチドであって、前記タンパク質にとって異種のシグナルペプチドは、必要に応じて、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されており、ただし前記タンパク質はオキシドレダクターゼではない、前記タンパク質にとって異種のシグナルペプチド;
ii)前記タンパク質に相同なシグナルペプチドであって、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、前記タンパク質に相同なシグナルペプチド;および
iii)自然状態でシグナルペプチドの機能を有さない天然に存在するアミノ酸配列であって、必要に応じて少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、天然に存在するアミノ酸配列
からなる群から選択される、mRNAに関する。
続きを表示(約 15,000 文字)
【0002】
本発明はさらに、i)タンパク質;およびii)前記タンパク質にとって異種のシグナルペプチドをコードする核酸配列を含むmRNAであって、前記タンパク質にとって異種のシグナルペプチドは、脳由来神経栄養因子(BDNF)のシグナルペプチドであり、前記タンパク質は、オキシドレダクターゼではない、mRNAに関する。また本発明は、タンパク質およびシグナルペプチドをコードする核酸配列を含む転写単位または発現ベクターであって、シグナルペプチドのアミノ酸配列のN末端のアミノ酸1~9は、2を超える平均疎水性スコアを有し、シグナルペプチドは、
i)前記タンパク質にとって異種のシグナルペプチドであって、前記タンパク質にとって異種のシグナルペプチドは、必要に応じて、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されており、ただし前記タンパク質はオキシドレダクターゼではない、前記タンパク質にとって異種のシグナルペプチド;
ii)前記タンパク質に相同なシグナルペプチドであって、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、前記タンパク質に相同なシグナルペプチド;および
iii)自然状態でシグナルペプチドの機能を有さない天然に存在するアミノ酸配列であって、必要に応じて少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、天然に存在するアミノ酸配列
からなる群から選択される、転写単位または発現ベクターにも関する。また本発明は、前記タンパク質にとって異種のタンパク質およびシグナルペプチドをコードする核酸配列を含む転写単位または発現ベクターであって、前記タンパク質にとって異種のシグナルペプチドは、脳由来神経栄養因子(BDNF)のシグナルペプチドである、転写単位または発現ベクターにも関する。また本発明は、mRNA、および/または転写単位もしくは発現ベクターを含む、治療用組成物およびキットにも関する。また本発明は、医薬として使用するための、mRNA、転写単位または発現ベクター、治療用組成物および/またはキットにも関し、特に、医薬として使用するための、i)IGF1;およびii)脳由来神経栄養因子(BDNF)のシグナルペプチドをコードする核酸配列を含むmRNAにも関する。また本発明は、骨格筋損傷を処置する方法での使用のためのmRNAおよびその治療用組成物にも関する。
【背景技術】
【0003】
これまでに、コードされたタンパク質の発現および分泌の収量を増加させるために、様々な試みが、特にインビトロおよび/またはインビボの両方での改善された発現系の使用によって、なされてきた。一般的に先行技術に記載された発現および分泌を増加させるための方法は、慣習的に、特異的なプロモーターおよび対応する調節エレメントを含有する発現ベクターまたはカセットの使用に基づく。これらの発現ベクターまたはカセットは、典型的には特定の細胞系に限定されるため、これらの発現系は、異なる細胞系で使用する場合、適合させる必要がある。次いでこのような適合させた発現ベクターまたはカセットは通常、細胞にトランスフェクトされ、典型的には特定の細胞株に応じて処置される。それ故に、主として、特定の細胞型に特異的なプロモーターおよび調節エレメントから独立した細胞に固有の系によって、標的細胞においてコードされたタンパク質を発現することができる、mRNAのような核酸分子が選ばれる。この状況において、mRNAを安定化するエレメントと、mRNAの翻訳効率を増加させるエレメントを区別することができる。例えば、WO02/098443は、一般的な形態で安定化され、そのコード化領域において翻訳に最適化されたmRNAを記載している。WO02/098443はさらに、配列修飾を決定するための方法を開示している。WO02/098443は、加えて、配列のグアニン/シトシン(G/C)含有量を増加させるためにmRNA配列中のアデニンおよびウラシルヌクレオチドを置換する可能性を記載している。この状況において、WO02/098443は、概して、このような修飾に関する塩基配列としての配列に言及しており、修飾されたmRNAは、処置しようとする患者で翻訳される少なくとも1つの生物学的に活性なペプチドまたはポリペプチドを、例えば、間違いがまったくないか、または不十分に、または多少の間違いを含んでのいずれかでコードする。コードされたタンパク質の発現を増加させるためのさらなるアプローチにおいて、WO2007/036366号出願は、長いポリ(A)配列(特に120bpより長い)ならびにmRNA安定性および翻訳活性へのベータグロビン遺伝子の少なくとも2つの3’非翻訳領域の組合せのプラスの作用を記載している。当業界における全ての進歩にもかかわらず、無細胞の系、細胞または生物体(組換え発現)におけるコードされたタンパク質の効率的な発現、特に効率的な分泌は、それでもなお挑戦しがいのある課題である。
【発明の概要】
【0004】
本発明は、タンパク質およびシグナルペプチドをコードする核酸配列を含むmRNAであって、シグナルペプチドのアミノ酸配列のN末端のアミノ酸1~9は、2を超える平均疎水性スコアを有し、シグナルペプチドは、
i)前記タンパク質にとって異種のシグナルペプチドであって、前記タンパク質にとって異種のシグナルペプチドは、必要に応じて、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されており、ただし前記タンパク質はオキシドレダクターゼではない、前記タンパク質にとって異種のシグナルペプチド;
ii)前記タンパク質に相同なシグナルペプチドであって、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、前記タンパク質に相同なシグナルペプチド;および
iii)自然状態でシグナルペプチドの機能を有さない天然に存在するアミノ酸配列であって、必要に応じて少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、天然に存在するアミノ酸配列
からなる群から選択される、mRNAを提供する。
【0005】
本発明はさらに、
i)タンパク質;および
ii)前記タンパク質にとって異種のシグナルペプチド
をコードする核酸配列を含むmRNAであって、
前記タンパク質にとって異種のシグナルペプチドは、脳由来神経栄養因子(BDNF)のシグナルペプチドであり、前記タンパク質は、オキシドレダクターゼではない、mRNAを提供し、特に、本発明は、
i)タンパク質;および
ii)前記タンパク質にとって異種のシグナルペプチド
をコードする核酸配列を含むmRNAであって、
前記タンパク質にとって異種のシグナルペプチドは、脳由来神経栄養因子(BDNF)のシグナルペプチドであり、前記タンパク質は、カルボキシペプチダーゼ;サイトカイン;細胞外のリガンドおよび輸送体;細胞外マトリックスタンパク質;グルコシダーゼ;グリコシルトランスフェラーゼ;増殖因子;増殖因子結合タンパク質;ヘパリン結合タンパク質;ホルモン;ヒドロラーゼ;免疫グロブリン;イソメラーゼ;キナーゼ;リアーゼ;金属酵素阻害剤;メタロプロテアーゼ;乳タンパク質;神経刺激性タンパク質;プロテアーゼ;プロテアーゼ阻害剤;タンパク質ホスファターゼ;エステラーゼ;トランスフェラーゼ;および血管作動性タンパク質からなる群から選択される、mRNAを提供する。
【0006】
本発明はさらに、タンパク質およびシグナルペプチドをコードする核酸配列を含む転写単位または発現ベクターであって、シグナルペプチドのアミノ酸配列のN末端のアミノ酸1~9は、2を超える平均疎水性スコアを有し、シグナルペプチドは、
i)前記タンパク質にとって異種のシグナルペプチドであって、前記タンパク質にとって異種のシグナルペプチドは、必要に応じて、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されており、ただし前記タンパク質はオキシドレダクターゼではない、前記タンパク質にとって異種のシグナルペプチド;
ii)前記タンパク質に相同なシグナルペプチドであって、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、前記タンパク質に相同なシグナルペプチド;および
iii)自然状態でシグナルペプチドの機能を有さない天然に存在するアミノ酸配列であって、必要に応じて少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、天然に存在するアミノ酸配列
からなる群から選択される、転写単位または発現ベクターを提供する。
【0007】
本発明はさらに、前記タンパク質にとって異種のシグナルペプチドは、脳由来神経栄養因子(BDNF)のシグナルペプチドであり、前記タンパク質は、オキシドレダクターゼではない、前記タンパク質にとって異種のタンパク質およびシグナルペプチドをコードする核酸配列を含む転写単位または発現ベクターを提供する。本発明はさらに、前記タンパク質にとって異種のシグナルペプチドは、脳由来神経栄養因子(BDNF)のシグナルペプチドであり、タンパク質は、カルボキシペプチダーゼ;サイトカイン;細胞外のリガンドおよび輸送体;細胞外マトリックスタンパク質;グルコシダーゼ;グリコシルトランスフェラーゼ;増殖因子;増殖因子結合タンパク質;ヘパリン結合タンパク質;ホルモン;ヒドロラーゼ;免疫グロブリン;イソメラーゼ;キナーゼ;リアーゼ;金属酵素阻害剤;メタロプロテアーゼ;乳タンパク質;神経刺激性タンパク質;プロテアーゼ;プロテアーゼ阻害剤;タンパク質ホスファターゼ;エステラーゼ;トランスフェラーゼ;および血管作動性タンパク質からなる群から選択される、前記タンパク質にとって異種のタンパク質およびシグナルペプチドをコードする核酸配列を含む転写単位または発現ベクターを提供する。本発明はさらに、上述のmRNAおよび/または転写単位もしくは発現ベクターを含む治療用組成物を提供する。本発明はさらに、上述のmRNA、転写単位または発現ベクターおよび/または治療用組成物、ならびに使用説明書、必要に応じてベクターマップ、必要に応じて宿主細胞、必要に応じて宿主細胞の培養のための培地、および/もしくは必要に応じてトランスフェクトされた宿主細胞を選択および培養するための選択培地を含むキットを提供する。本発明はさらに、医薬として使用するための、上述のmRNA、転写単位または発現ベクター、治療用組成物またはキットを提供する。本発明はさらに、医薬として使用するための、i)IGF1;およびii)脳由来神経栄養因子(BDNF)のシグナルペプチドをコードする核酸配列を含むmRNAを提供する。本発明はさらに、骨格筋損傷を処置する方法での使用のための、mRNAまたはmRNAを含有する治療用組成物を提供する。
【0008】
本発明者らは、驚くべきことに、タンパク質およびシグナルペプチドをコードする核酸配列を含むmRNAであって、シグナルペプチドのアミノ酸配列のN末端のアミノ酸1~9は、2を超える平均疎水性スコアを有し、シグナルペプチドは、
i)前記タンパク質にとって異種のシグナルペプチドであって、前記タンパク質にとって異種のシグナルペプチドは、必要に応じて、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されており、ただし前記タンパク質はオキシドレダクターゼではない、前記タンパク質にとって異種のシグナルペプチド;
ii)前記タンパク質に相同なシグナルペプチドであって、少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、前記タンパク質に相同なシグナルペプチド;および
iii)自然状態でシグナルペプチドの機能を有さない天然に存在するアミノ酸配列であって、必要に応じて少なくとも1つのアミノ酸の挿入、欠失および/または置換によって修飾されている、天然に存在するアミノ酸配列
からなる群から選択される、mRNAは、タンパク質およびその天然の相同なシグナルペプチドをコードするmRNAでトランスフェクトされた細胞によるタンパク質の分泌と比較して、このmRNAでトランスフェクトされた細胞によるタンパク質のより効率的な分泌を提供することを見出した。特に、本発明者らは、驚くべきことに、i)タンパク質;およびii)前記タンパク質にとって異種のBDNFシグナルペプチドをコードする核酸配列を含むmRNAは、タンパク質の天然の相同なシグナルペプチドと比べて、このmRNAでトランスフェクトされた細胞によるタンパク質のより効率的な分泌を提供することを見出した。分泌タンパク質の量は、同じタンパク質およびタンパク質の天然の相同なシグナルペプチドを含むmRNAと比較して、最大で6倍多い。この予想外の発見は、所望のタンパク質をコードするmRNAを効果的に送達し、細胞中で発現させ、タンパク質の天然の相同なシグナルペプチドを有するものより多くの量の分泌タンパク質を得るのに非常に有用である。本発明によって提供される同じ量のmRNAを用いて得られた、より多くの量の分泌タンパク質は、治療用量を、組織に局所適用するのに必要な用量より少なくし、それによって起こり得るmRNA関連の副作用に対するその安全性の枠を増加させるために極めて有用である。さらにこれは、本出願を、制御放出のための製剤やデバイスのコーティングにより適したものにしている。さらにこれは、mRNA関連の免疫原性のリスクを低減し、本出願を、限られた体積しか注射できない組織に対して、またはこれまで処理しにくい組織により適したものにしている。本発明者らはまた、mRNA、特にヒトIGF-1をコードするmRNAを骨格筋に効果的に送達し、発現させることを可能にし、それによって骨格筋における所望のポリペプチドの発現を可能にし、意味のある機能的な利益が筋肉に提供されることも見出した。mRNAは、好ましくは液体組成物で、好ましくは裸の形態で存在する。この液体組成物は、例えば注射によって骨格筋に直接送達することができ、mRNAのための遺伝子移入ベクターまたは担体、またはエレクトロトランスファーまたは超音波のような組織への移入を強化するための方法をまったく必要としない。さらに、損傷を受けた骨格筋へのmRNAの注射は、回復プロセスを促進し、骨格筋の機能の増加をもたらすことが示された。驚くべきことに、IGF-1をコードするmRNAで処置した動物は、16日間の間に(by 16 days)健康な範囲の機能的なレベルに到達した。対照的に、ビヒクルで処置した対照動物は、28日目でさえも十分な機能的な回復を達成しなかった。
【図面の簡単な説明】
【0009】
Cpd.1のDNAおよびRNA配列を示す図である。(A)は、そのプレドメイン、プロドメインおよびコードドメインを含有するヒトコドン最適化IGF1のDNA配列(配列番号1)を示す。プレドメイン(シグナル伝達ペプチド)の配列はイタリック体で示され、プロドメインの配列は
下線が引かれ
、IGF-Iコードドメインは太字で示され、ストップコドンは
太字
で示されている。(B)は、ウリジンがN1-メチルシュードウリジンである、ヒトIGF1のプレドメイン、プロドメインおよびコードドメインのRNA配列を図示する(配列番号2)。プレドメインおよびプロドメインは分泌時に切断される。
Cpd.2のDNAおよびRNA配列を示す図である。(A)は、IGF2プレドメインならびにIGF1プロドメインおよびコードドメインを含有するヒトコドン最適化IGF1のDNA配列(配列番号3)を示す。プレドメイン(シグナル伝達ペプチド)の配列はイタリック体で示され、プロドメインの配列は
下線が引かれ
、IGF1コードドメインは太字で示され、ストップコドンは
太字
で示されている。(B)は、ウリジンがN1-メチルシュードウリジンである、IGF2プレドメインならびにIGF1プロドメインおよびコードドメインのRNA配列を図示する(配列番号4)。プレドメインおよびプロドメインは分泌時に切断される。
Cpd.3のDNAおよびRNA配列を示す図である。(A)は、ALBプレドメインならびにIGF1プロドメインおよびコードドメインを含有するヒトコドン最適化IGF1のDNA配列(配列番号5)を示す。プレドメイン(シグナル伝達ペプチド)の配列はイタリック体で示され、プロドメインの配列は
下線が引かれ
、IGF1コードドメインは太字で示され、ストップコドンは
太字
で示されている。(B)は、ウリジンがN1-メチルシュードウリジンである、ALBプレドメインおよびIGF1プロドメインおよびコードドメインのRNA配列を図示する(配列番号6)。プレドメインおよびプロドメインは分泌時に切断される。
Cpd.4のDNAおよびRNA配列を示す図である。(A)は、BDNFプレドメインならびにIGF1プロドメインおよびコードドメインを含有するヒトコドン最適化IGF1のDNA配列(配列番号7)を示す。プレドメイン(シグナル伝達ペプチド)の配列はイタリック体で示され、プロドメインの配列は
下線が引かれ
、IGF1コードドメインは太字で示され、ストップコドンは
太字
で示されている。(B)は、ウリジンがN1-メチルシュードウリジンである、BDNFプレドメインならびにIGF1プロドメインおよびコードドメインのRNA配列を図示する(配列番号8)。プレドメインおよびプロドメインは分泌時に切断される。
Cpd.5のDNAおよびRNA配列を示す図である。(A)は、CXCL12プレドメインならびにIGF1プロドメインおよびコードドメインを含有するヒトコドン最適化IGF1のDNA配列(配列番号9)を示す。プレドメイン(シグナル伝達ペプチド)の配列はイタリック体で示され、プロドメインの配列は
下線が引かれ
、IGF1コードドメインは太字で示され、ストップコドンは
太字
で示されている。(B)は、ウリジンがN1-メチルシュードウリジンである、CXCL12プレドメインならびにIGF1プロドメインおよびコードドメインのRNA配列を図示する(配列番号10)。プレドメインおよびプロドメインは分泌時に切断される。
Cpd.6のDNAおよびRNA配列を示す図である。(A)は、合成シグナル伝達ペプチド1プレドメインならびにIGF1プロドメインおよびコードドメインを含有するヒトコドン最適化IGF1のDNA配列(配列番号11)を示す。プレドメイン(シグナル伝達ペプチド)の配列はイタリック体で示され、プロドメインの配列は
下線が引かれ
、IGF1コードドメインは太字で示され、ストップコドンは
太字
で示されている。(B)は、ウリジンがN1-メチルシュードウリジンである、合成シグナル伝達ペプチド1プレドメインならびにIGF1プロドメインおよびコードドメインのRNA配列を図示する(配列番号12)。プレドメインおよびプロドメインは分泌時に切断される。
Cpd.7のDNAおよびRNA配列を示す図である。(A)は、合成シグナル伝達ペプチド2プレドメインならびにIGF1プロドメインおよびコードドメインを含有するヒトコドン最適化IGF1のDNA配列(配列番号13)を示す。プレドメイン(シグナル伝達ペプチド)の配列はイタリック体で示され、プロドメインの配列は
下線が引かれ
、IGF1コードドメインは太字で示され、ストップコドンは
太字
で示されている。(B)は、ウリジンがN1-メチルシュードウリジンである、合成シグナル伝達ペプチド2プレドメインならびにIGF1プロドメインおよびコードドメインのRNA配列を図示する(配列番号14)。プレドメインおよびプロドメインは分泌時に切断される。
太字
でマークされたCpd.1インサートを含むベクターpVAX.A120のDNA配列を示す図である(配列番号15)。Cpd.1のORFは、その元のプラスミドから消化され、ベクターにサブクローニングされた。
太字
でマークされたCpd.2インサートを含むベクターpMA-TのDNA配列を示す図である(配列番号16)。Cpd.2のORFは、その元のプラスミドから消化され、ベクターにサブクローニングされた。
太字
でマークされたCpd.3インサートを含むベクターpMA-TのDNA配列を示す図である(配列番号17)。Cpd.3のORFは、その元のプラスミドから消化され、ベクターにサブクローニングされた。
太字
でマークされたCpd.4インサートを含むベクターpMA-TのDNA配列を示す図である(配列番号18)。Cpd.4のORFは、その元のプラスミドから消化され、ベクターにサブクローニングされた。
太字
でマークされたCpd.5インサートを含むベクターpMA-TのDNA配列を示す図である(配列番号19)。Cpd.5のORFは、その元のプラスミドから消化され、ベクターにサブクローニングされた。
太字
でマークされたCpd.6を含むベクターpMA-RQのDNA配列を示す図である(配列番号20)。Cpd.6のORFは、その元のプラスミドから消化され、ベクターにサブクローニングされた。
太字
でマークされたCpd.7を含むベクターpMA-RQのDNA配列を示す図である(配列番号21)。Cpd.7のORFは、その元のプラスミドから消化され、ベクターにサブクローニングされた。
mRNAのIVT用pMA-TおよびpMA-RQプラスミドを増幅するのに使用されるフォワード(配列番号22)およびリバースプライマー(配列番号23)配列を示す図である。
Cpd.1~Cpd.7のシグナルペプチドの遺伝子名、UniProt数、コドン最適化DNA配列およびアミノ酸配列ならびにベクター(配列番号24~37)を示す図である。Cpd.6およびCpd.7のシグナルペプチドは合成ペプチドであり、公開データベース中の一致する既知のタンパク質配列ではないことに注意。
Cpd.1~Cpd.7のmRNAトランスフェクションによる、ヒト胎児性腎細胞(HEK293T)からのIGF1分泌の誘導を示す図である。HEK293T細胞は各2μg Cp.1~Cpd.7をトランスフェクトされ、分泌されたIGF1は、特異的ELISAを使用して24時間後に細胞培養上清で測定された。Cpd.4は、Cpd.1より有意に高いIGF1分泌を誘導した(3.3倍)。データは、4つの複製物の平均±平均の標準誤差を表す。有意性(
***
、<0.001)は、一方向ANOVAとその後のダネットの多重比較検定によって評価された。
Cpd.1またはCpd.4のmRNAトランスフェクション後のHEK293T細胞でのIGF1分泌の誘導の濃度依存性を示す図である。細胞は、種々の濃度(0、0.02、0.06、0.2、0.6または2μg)でCpd.1またはCpd.4をトランスフェクトされ、分泌されたIGF1は、特異的ELISAを使用して24時間後に細胞培養上清で測定された。Cpd.4誘導IGF1分泌は、Cpd.1(EC
50
0.889μg)より有意に強力であった(EC
50
0.134μg)。データは、2つの複製物の平均±平均の標準誤差を表す。有意性(
***
、<0.001)は、2つの曲線の二方向ANOVAによって評価された。
Cpd.1~Cpd.7のmRNAトランスフェクションによる、マウス骨格筋細胞(C2C12)からのIGF1分泌の誘導を示す図である。C2C12細胞は各2μg Cp.1~Cpd.7をトランスフェクトされ、分泌されたIGF1は、特異的ELISAを使用して24時間後に細胞培養上清で測定された。Cpd.4は、Cpd.1より有意に高いIGF1分泌を誘導した(6.1倍)。データは、4つの複製物の平均±平均の標準誤差を表す。有意性(
***
、<0.001)は、一方向ANOVAとその後のダネットの多重比較検定によって評価された。
Cpd.1およびCpd.4のmRNAトランスフェクションによる、ヒト初代骨格筋細胞(HSkMC)からのIGF1分泌の誘導を示す図である。HSkMC細胞は各2μg Cp.1またはCpd.4をトランスフェクトされ、分泌されたIGF1は、特異的ELISAを使用して24時間後に細胞培養上清で測定された。Cpd.4は、Cpd.1より有意に高いIGF1分泌を誘導した(3.1倍)。データは、3つの複製物の平均±平均の標準誤差を表す。有意性(
**
、p<0.01)は、一方向ANOVAとその後のダネットの多重比較検定によって評価された。
ノテキシン損傷後の前脛骨筋(TA)の機能回復を示す図である。筋肉内注射(0日目)によるノテキシン損傷後、2つのIGF-I mRNA処置(Cpd.4(1μg))が1日目および4日目の筋肉内注射によって適用された(矢印の頭を参照)。対照群はビヒクル溶液を受け取った。筋機能は、損傷後1、4、7、10、14、21および28日目に評価された。データは、1群および1時点あたり5マウスの平均±平均の標準誤差(SEM)を表す。アスタリスクは、スチューデントのt検定によって評価された場合のCpd.4処置群と対照群との有意差(p<0.05)を示す。
対照としてのCpd.1およびCpd.8~Cpd.26のmRNAトランスフェクションによる、ヒト胎児性腎細胞(HEK293T)からのIGF1分泌の誘導を示す図である。HEK293T細胞は各0.3μg Cpd.1およびCpd.8~Cpd.26をトランスフェクトされ、分泌されたIGF1は、特異的ELISAを使用して24時間後に細胞培養上清で測定された。IGF1分泌はCpd.1 IGF1分泌に対して正規化された。Cpd.8、9、10、11、12および13はIGF1分泌の低下を示したのに対し、Cpd.14、15、16、17、18、19、20、21、23、24、25および26はCpd.1より高いIGF1分泌を誘導した(最大2.6倍)。データは、1Cpd.につき2~11の複製物の平均±平均の標準誤差を表す。有意性(
*
、p<0.05;
**
、p<0.001;
***
、<0.001)は、Cpd.1と比較した個々のCpd.のスチューデントのt検定によって評価された。
対照としてのCpd.1およびCpd.4~Cpd.26のmRNAトランスフェクションによる、ヒト肝細胞(HepG2)からのIGF1分泌の誘導を示す図である。HepG2細胞は各0.3μg Cp.1およびCpd.4~Cpd.26をトランスフェクトされ、分泌されたIGF1は、特異的ELISAを使用して24時間後に細胞培養上清で測定された。IGF1分泌はCpd.1に対して正規化された。Cpd.8、9および12はIGF1分泌の低下を示したのに対し、Cpd.4、15、16、17、18、19、20、21、22、23、24、25および26はCpd.1より高いIGF1分泌を誘導した(最大8.3倍)。データは、1Cpd.につき2~4つの複製物の平均±平均の標準誤差を表す。有意性(
**
、p<0.01;
***
、<0.001)は、Cpd.1と比較した個々のCpd.のスチューデントのt検定によって評価された。
対照としてのCpd.1およびCpd.4~Cpd.24のmRNAトランスフェクションによる、ヒト神経細胞(IMR32)からのIGF1分泌の誘導を示す図である。IMR32細胞は各0.3μg Cp.1およびCpd.4~Cpd.24をトランスフェクトされ、分泌されたIGF1は、特異的ELISAを使用して24時間後に細胞培養上清で測定された。IGF1分泌はCpd.1に対して正規化された。Cpd.4、14、15、16、17、20、22、23および24はCpd.1より高いIGF1分泌を誘導した(最大2.6倍)。データは、1Cpd.につき2~6つの複製物の平均±平均の標準誤差を表す。有意性(
*
、p<0.05;
***
、<0.001)は、Cpd.1と比較した個々のCpd.のスチューデントのt検定によって評価された。
対照としてのCpd.1およびCpd.4~Cpd.25のmRNAトランスフェクションによる、ヒト初代軟骨細胞からのIGF1分泌の誘導を示す図である。軟骨細胞は各0.6μg Cp.1およびCpd.4~Cpd.25をトランスフェクトされ、分泌されたIGF1は、特異的ELISAを使用して24時間後に細胞培養上清で測定された。IGF1分泌はCpd.1に対して正規化された。Cpd.4、14、15、16、20、21、22、24および25はCpd.1より高いIGF1分泌を誘導した(最大1.9倍)。データは、1Cpd.につき1~2つの複製物の平均±平均の標準誤差を表す。有意性(
*
、p<0.05;
***
、<0.001)は、Cpd.1と比較した個々のCpd.のスチューデントのt検定によって評価された。
対照としてのCpd.1およびCpd.4~Cpd.17のmRNAトランスフェクションによる、ラット野生型(A)またはSOD1G
S93A
(B)初代運動ニューロンからのIGF1分泌の誘導を示す図である。ラット野生型初代運動ニューロンは、各0.3μg Cp.1、Cpd.4、Cpd.14およびCpd.17をトランスフェクトされ、ラットSOD1G
S93A
初代運動ニューロンは、各0.3μg Cp.1、Cpd.14およびCpd.17をトランスフェクトされ、分泌されたIGF1は、特異的ELISAを使用して48時間後に細胞培養上清で測定された。IGF1分泌はCpd.1に対して正規化された。Cpd.4、14および17はCpd.1より高いIGF1分泌を誘導した(野生型で最大4.3倍またはSOD1
S93A
で9.3倍)。データは、1Cpd.につき2つの複製物の平均±平均の標準誤差を表す。有意性は、Cpd.1と比較した個々のCpd.のスチューデントのt検定によって評価され、統計的差異がないことを明らかにした。
Cpd.27、Cpd.28またはCpd.29のmRNAトランスフェクションによる、ヒト胎児性腎細胞(HEK293T、A)、ヒト肝細胞(HepG2、B)およびヒト肺癌細胞(A549、C)からのEPO分泌の誘導を示す図である。細胞は各0.3~0.9μg Cp.27、Cpd.28またはCpd.29をトランスフェクトされ、分泌されたEPOは、特異的ELISAを使用して24時間後に細胞培養上清で測定された。EPO分泌はCpd.27に対して正規化された。Cpd.28および29は、分析された3つの細胞型全てにおいてCpd.27より高いEPO分泌を誘導した(最大1.8倍)。データは、1Cpd.につき3~8つの複製物の平均±平均の標準誤差を表す。有意性(
*
、p<0.05;
***
、<0.001)は、Cpd.27と比較した個々のCpd.のスチューデントのt検定によって評価された。
Cpd.30、Cpd.31またはCpd.32のmRNAトランスフェクションによる、ヒト胎児性腎細胞(HEK293T)からのINS分泌の誘導を示す図である。細胞は各0.6μg Cp.30、Cpd.31またはCpd.32をトランスフェクトされ、分泌されたINSは、特異的ELISAを使用して24時間後に細胞培養上清で測定された。INS分泌はCpd.30に対して正規化された。Cpd.31および32はCpd.30より高いINS分泌を誘導した(最大3.9倍)。データは、1Cpd.につき3~5つの複製物の平均±平均の標準誤差を表す。有意性(
*
、p<0.05;
***
、<0.001)は、Cpd.30と比較した個々のCpd.のスチューデントのt検定によって評価された。
Cpd.33、Cpd.34またはCpd.35のmRNAトランスフェクションによる、ヒト胎児性腎細胞(HEK293T、A)、ヒト肝細胞(HepG2、B)、ヒト単球(THP-1、C)およびヒト肺癌細胞(A549、D)からのIL4分泌の誘導を示す図である。細胞は各0.5~0.6μg Cp.33、Cpd.34またはCpd.35をトランスフェクトされ、分泌されたIL4は、特異的ELISAを使用して24時間後に細胞培養上清で測定された。IL4分泌はCpd.33に対して正規化された。Cpd.34および35は、分析された3つの細胞型全てにおいてCpd.33より高いIL4分泌を誘導した。in(最大2.2倍)。データは、1Cpd.につき3~8つの複製物の平均±平均の標準誤差を表す。有意性(
*
、p<0.05;
***
、<0.001)は、Cpd.33と比較した個々のCpd.のスチューデントのt検定によって評価された。
Cpd.36、Cpd.37またはCpd.38のmRNAトランスフェクションによる、ヒト胎児性腎細胞(HEK293T、A)、ヒト肝細胞(HepG2、B)またはヒト単球(THP-1、C)からのIL10分泌の誘導を示す図である。細胞は各0.3~0.6μg Cp.36、Cpd.37またはCpd.38をトランスフェクトされ、分泌されたIL10は、特異的ELISAを使用して24時間後に細胞培養上清で測定された。IL10分泌はCpd.36に対して正規化された。Cpd.37および38は、分析された3つの細胞型全てにおいてCpd.36より高いIL10分泌を誘導した(最大2.2倍)。データは、1Cpd.につき4~8つの複製物の平均±平均の標準誤差を表す。有意性(
**
、p<0.01;
***
、<0.001)は、Cpd.36と比較した個々のCpd.のスチューデントのt検定によって評価された。
Cpd.39のmRNAトランスフェクションによる、ヒト肝細胞(HepG2、A)およびヒト初代軟骨細胞(B)からのIGF-1分泌の誘導を示す図である。細胞は各0.3~0.6μg Cp.39をトランスフェクトされ、分泌IGF1は、特異的ELISAを使用して24時間後に細胞培養上清で測定された。IGF1分泌はCpd.1に対して正規化された。Cpd.39は、分析された2つの細胞型全てにおいてCpd.1より高いIGF1分泌を誘導した(最大1.4倍)。データは、4~7つの複製物の平均±平均の標準誤差を表す。有意性(
**
、p<0.01;
***
、<0.001)は、Cpd.39と比較した個々のCpd.1のスチューデントのt検定によって評価された。
【発明を実施するための形態】
【0010】
用語「RNA」は、本明細書で使用される場合、アミノ酸配列をコードするRNA、加えて、アミノ酸配列をコードしないRNAを含む。通常、本明細書で使用されるようなRNAは、コードRNAであり、すなわちアミノ酸配列をコードするRNAである。このようなRNA分子はまた、mRNA(メッセンジャーRNA)とも称され、一本鎖RNA分子である。したがって、用語「RNA」は、本明細書で使用される場合、好ましくはmRNAを指す。RNAは、当業者公知の化学的および酵素的な合成手法によって、または組換え技術の使用によって作製してもよいし、または天然源から単離してもよいし、またはそれらの組合せであってもよい。RNAは、必要に応じて、天然にはない、および天然に存在するヌクレオシド修飾、例えば、本明細書ではメチルプソイドウリジンとも称されるN
1
-メチルプソイドウリジンなどを含んでいてもよい。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
個人
酒類
28日前
株式会社オシキリ
発酵検査装置
17日前
松谷化学工業株式会社
澱粉分解物の製造方法
15日前
東ソー株式会社
ポリマーコートビーズの製造方法
16日前
インヒブルクス バイオサイエンシズ インコーポレイテッド
CLEC12a結合性ポリペプチド及びその使用
1か月前
本田技研工業株式会社
培養システム
1か月前
豊田合成株式会社
細胞培養膜及び細胞培養方法
1か月前
東ソー株式会社
アデノ随伴ウイルスのスクリーニング方法
28日前
住友化学株式会社
積層体
1か月前
三井化学株式会社
培養容器
23日前
株式会社渡辺オイスター研究所
脂肪細胞の成長抑制剤
16日前
住友化学株式会社
細胞チップ
1か月前
旭化成ファーマ株式会社
ジアホラーゼ
29日前
ZACROS株式会社
培養装置、及び培養装置積層体
28日前
株式会社トクヤマ
乾燥用保護剤、生体試料測定試薬、及び分析方法
17日前
公立大学法人宮城大学
D-セリンの製造方法
15日前
ウシオ電機株式会社
培養容器
29日前
雪印メグミルク株式会社
D-セリン濃度低減用組成物
15日前
グリーンケミカルズ株式会社
フェノール組成物およびポリマー
10日前
学校法人近畿大学
新規酵母と柿を用いた酒類の製造方法
1か月前
ニッタ株式会社
凍結細胞解凍装置
15日前
サントリーホールディングス株式会社
ビールテイスト飲料
28日前
学校法人東邦大学
自己反応性T細胞の検出方法
9日前
東ソー株式会社
細胞外小胞の解析方法
15日前
株式会社ジャパンディスプレイ
検出装置
29日前
株式会社ジャパンディスプレイ
検出装置
29日前
サッポロビール株式会社
発酵ビールテイストアルコール飲料
1か月前
理研食品株式会社
マツモの付着器の培養方法
8日前
アサヒビール株式会社
ビール様飲料の静菌性を増強する方法
8日前
日産化学株式会社
無血清培地中での細胞培養用下地材料
8日前
TOPPANホールディングス株式会社
コロニー計数システム
1か月前
国立大学法人 奈良先端科学技術大学院大学
胃幹細胞培養用培地、培養方法
8日前
高須合資会社
遠心分離による日本酒製造方法
15日前
東ソー株式会社
耐アルカリ性の向上したアデノ随伴ウイルス結合性タンパク質
29日前
株式会社日立ハイテク
自動培養装置、自動培養システム
22日前
国立大学法人 東京医科歯科大学
肝細胞オルガノイド及びその製造方法
22日前
続きを見る
他の特許を見る





 特許ウォッチ
特許ウォッチ