TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2024095710
公報種別
公開特許公報(A)
公開日
2024-07-10
出願番号
2024050953,2021169192
出願日
2024-03-27,2017-05-05
発明の名称
胃底部組織のインビトロでの製造のための方法及び当該方法と関連した組成物
出願人
チルドレンズ ホスピタル メディカル センター
代理人
個人
主分類
C12N
5/071 20100101AFI20240703BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】ヒトの胃の胃底部上皮を研究するための適切なモデルであるヒト胃底部オルガノイド(hFGO)を提供する。
【解決手段】a)MUC5AC陽性表面粘膜細胞及び/又はMUC6陽性粘膜頚細胞と、b)グレリン(GHRL)、ソマトスタチン(SST)、及び/又はヒスタミンを発現する内分泌細胞と、c)MIST1、ペプシノーゲンA(PGA5)、及び/又はペプシノーゲンC(PGC)を発現する主細胞と、d)傍細胞の密な腺と、及び/又はe)ATP4A、ATP4B、及びGIFを発現する傍細胞と、を特徴とする、ヒト胃底部オルガノイド(hFGO)とする。
【選択図】なし
特許請求の範囲
【請求項1】
胃底部組織の形成を誘導するインビトロ方法であって、
a)哺乳類胚体内胚葉(DE)細胞を、wnt経路活性化因子、FGFシグナル伝達経路活性化因子、BMPシグナル伝達経路阻害因子、及びレチノイン酸と第1の期間接触させるステップであって、
前記第1の期間は、前記胚体内胚葉から3次元後部前腸スフェロイドを形成するのに十分な長さの期間である、ステップと、
b)前記3次元後部前腸スフェロイドを、基底膜マトリックス中で成長因子、前記Wntシグナル伝達経路活性化因子、前記EGFシグナル伝達経路活性化因子、前記BMPシグナル伝達経路阻害因子、及びレチノイン酸とともに第2の期間懸濁させるステップであって、前記第2の期間は、胃底部hGO(hFGO)を含む胃底部系譜を誘導するのに十分な長さの期間である、ステップと、
c)ステップb)の前記hFGOを前記wnt経路活性化因子及び前記EGFシグナル伝達経路活性化因子とともに第3の期間培養するステップと、
d)ステップcの前記hFGOを前記wntシグナル伝達経路活性化因子、前記EGFシグナル伝達経路活性化因子、及びFGF10とともに第4の期間培養するステップと、
e)ステップdの前記hFGOをMEK阻害因子と第5の期間接触させるステップであって、前記第5の期間は、機能的胃底部細胞タイプを含む前記胃底部組織を形成するのに十分な期間である、ステップと、を含む、方法。
続きを表示(約 570 文字)
【請求項2】
前記第1の期間は、3日間±24時間であり、前記レチノイン酸は、前記期間±24時間の3日目に添加される、請求項1に記載の方法。
【請求項3】
前記第2の期間は、3日間±24時間である、請求項1に記載の方法。
【請求項4】
前記第3の期間は、11日間±24時間である、請求項1に記載の方法。
【請求項5】
前記第4の期間は、10日間±24時間である、請求項1に記載の方法。
【請求項6】
前記第5の期間は、2日間の期間±24時間である、請求項1に記載の方法。
【請求項7】
ステップe)は、前記胃底部hGOをBMP4シグナル伝達の活性化因子と接触させるステップをさらに含む、請求項1に記載の方法。
【請求項8】
前記機能的胃底部細胞タイプは、プロトンポンプタンパク質を発現し、かつ酸を分泌する傍細胞である、請求項1に記載の方法。
【請求項9】
前記機能的胃底部細胞タイプは、ペプシノーゲンを分泌する主細胞である、請求項1に記載の方法。
【請求項10】
前記ステップeは、SOX2+GATA+PDX1上皮を発達させるのに十分な期間実施される、請求項1に記載の方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
関連出願の相互参照
本出願は、2016年5月5日出願の米国仮特許出願第62/332,194号に対する優先権及び利益を主張し、その内容がすべて参照により組み込まれる。
続きを表示(約 8,500 文字)
【0002】
政府支援条項
本発明は、All 16491及びDK092456の下での政府の支援を用いて行われた。当該政府は、本発明におけるある特定の権利を有する。
【背景技術】
【0003】
胃疾患が地球規模で蔓延しているにもかかわらず、ヒトの胃の胃底部上皮を研究するための適切なモデルは少数である。ヒト胃底部型胃オルガノイド(hFGO)の開発は、ヒトの胃の生理学、病理生理学、及び薬剤開発の分子レベルの基礎を研究するための新規のかつ強力なモデルシステムであろう。
【発明の概要】
【0004】
本開示は、哺乳類胚体内胚葉(DE)細胞を特定の組織(複数可)または器官(複数可)へと、定方向分化を通じて転換するための方法に関する。特に、本開示は、分化した胚体内胚葉から形成される胃底部組織及び/またはオルガノイドの形成に関する。
【図面の簡単な説明】
【0005】
当業者は、以下に説明する図面が、単に説明目的のためにのみあることを理解するであろう。図面は、いかなる方法においても本教示の範囲を限定するよう企図するものではない。
【0006】
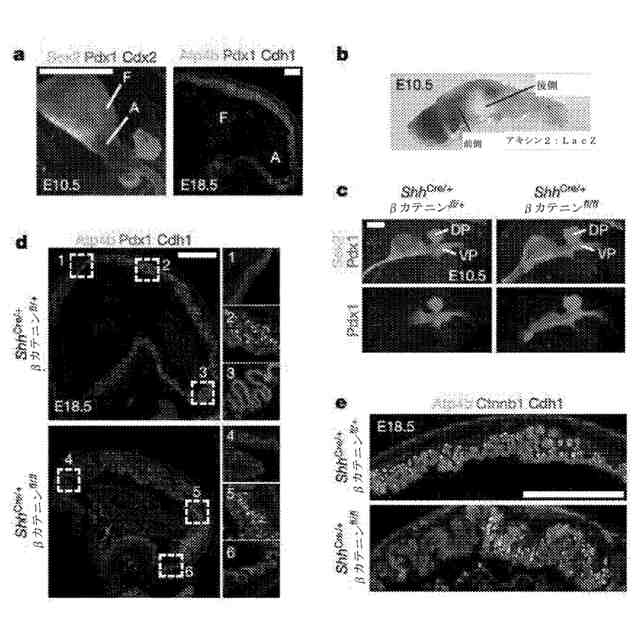
Wnt/βカテニンシグナル伝達は、マウスにおける胚性胃底部の特異性に必要である。a:Pdx1及びSox2を前庭(a)において発現させたのに対し、Pdx1は、18.5日胚でAtp4b発現傍細胞によって識別されるように、胃底部(f)に存在しなかった。b:Axin2:LacZレポーター胚由来の10.5日胚の前腸のX-gal染色は、Wnt活性が、胃の前部に制限されているが、後部胃からは除外されていることを示した。c:胃上皮におけるβカテニンの欠失は、胃の胃底部領域へのPdx1の前部増殖を生じた。d:18.5日のShh
Cre/+
βカテニン
fl/fl
(cKO)胚において、Pdx1を胃のいたるところに発現させたが、例外は、壁細胞含有上皮のいくらかの残余のパッチにおいてであった。挿絵1a~1c及び2a~2cはそれぞれ、対照の胃及びcKO胃における箱で囲まれた領域を示す。e、cKO胃において、Ctnnb1は、モザイク状欠失を呈し、壁細胞は、Ctnnb1の十分な上皮において分化しかしなかった。目盛尺:250μm(a)、200μm(c)、及び500μm(d及びe)。
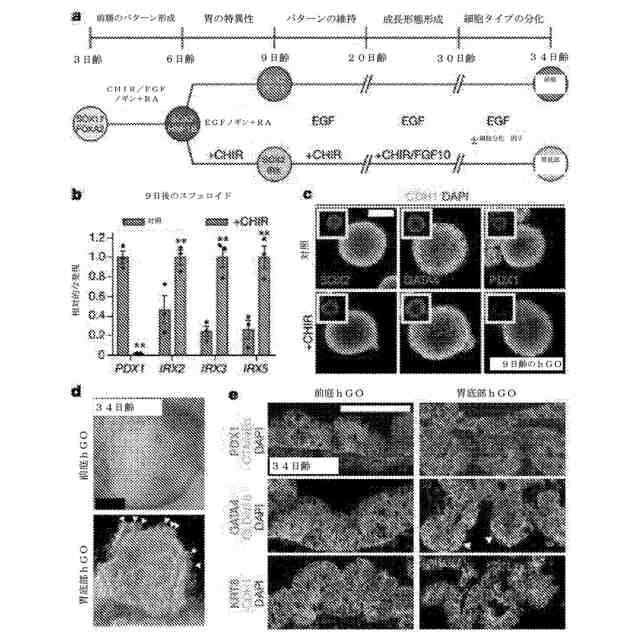
βカテニンの活性化は、ヒト前腸前駆体スフェロイドからの胃底部の発達を促進する。a:胃底部及び前庭の両方のhGOのための分化プロトコルの図式化した図。b、c:9日後に、CHIR処理したオルガノイドは、PDX1の減少、IRX2、IRX3及びIRX5の増加、ならびに胃マーカーSOX2またはGATA4の変化がないことを呈した。*:p<0.05、両側スチューデントt検定、n=3の生物学的複製物、データは4回の独立した実験を代表。d:hFGOは、hAGOに匹敵して生育しただけでなく、腺出芽形態形成(白色の矢頭)も呈した。e:両方のhGOは、CDH1、KRT8、及びCTNNB1ならびに胃マーカーGATA4及びCLDN18を発現する上皮を含有していた。hAGOは、ほぼ遍在的なPDX1発現を呈したのに対し、hFGOは、呈しなかった。目盛尺:50μm(c)、500μm(d)及び100μm(e)。エラーバーは、平均値の標準誤差を表す。
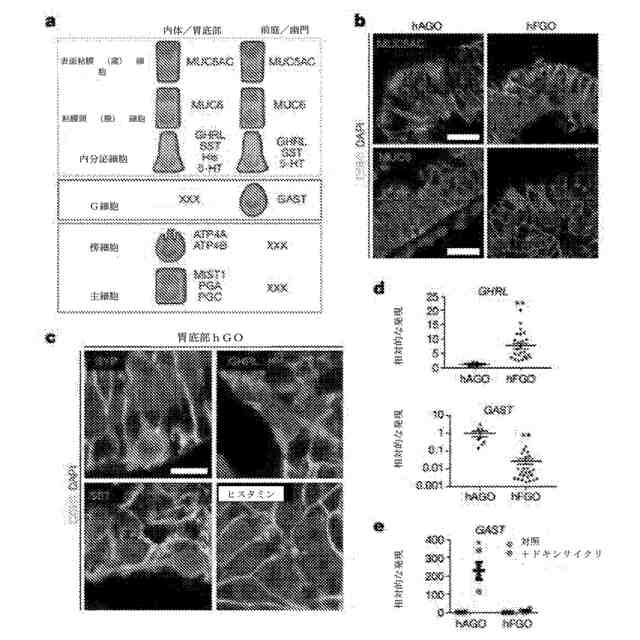
hGOにおける粘膜及び内分泌細胞系譜の分化。a:胃の胃底部及び前庭の腺において認められた共有されかつ異なる系譜の模式図。b:前庭及び胃底部のhGOは両方とも、MUC5AC陽性表面粘膜細胞及びMUC6陽性粘膜頸細胞を含有した。c、d:hFGOは、傍内分泌マーカーSYPを発現する内分泌細胞を含有した。異なるホルモン細胞タイプをhFGOにおいて識別し、これには、GHRL発現内分泌細胞、SST発現内分泌細胞、及びヒスタミン発現内分泌細胞を含んでいた。前庭特異的G細胞マーカーであるGASTは、hAGOでは発現したが、hFGOでは発現せず、逆に、GHRLは、hFGOにおいて多量であった。**:p<0.01、両側スチューデントt検定、hAGO及びhFGOにおいてそれぞれn=8及び24の生物学的複製物、データは6回の独立した実験を代表。e:hAGOは、プロ内分泌転写因子NEUROG3(+dox)の発現に応じて、前庭特異的GAST発現内分泌細胞を生じる能力があったが、hFGOはそうではなかった。*:p<0.01、両側スチューデントt検定、n=4の生物学的複製物、データは3回の独立した実験を代表。エラーバーは、平均値の標準誤差を表す。
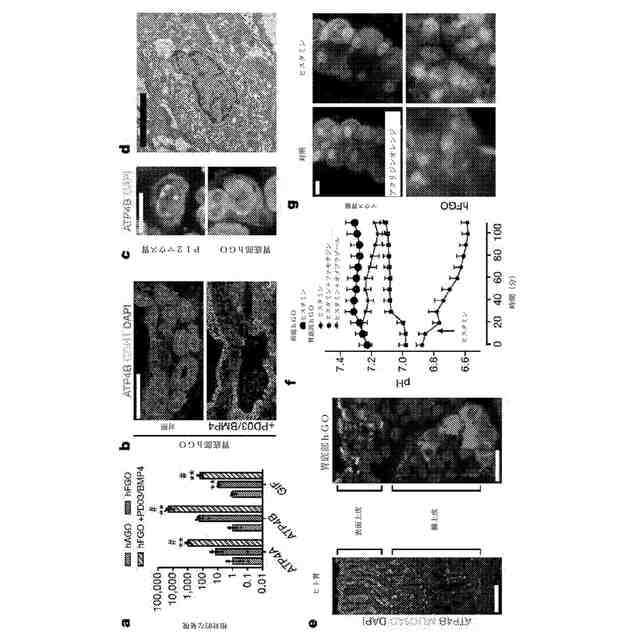
hFGOにおける主細胞の形成。a:hFGOは、MIST1及びペプシノーゲンC(PGC)の両方に陽性の細胞を有していた。b:尖端のPGC染色を有する一群の細胞を有する腺を示すパネル(a)における箱で囲まれた領域の高解像度。c:hFGOは、hAGOと比較して、主細胞マーカーPGA5(1000倍)、PGC(100倍)、及びMIST1(10倍超)の有意に高い発現を有していた。**:p<0.05、両側スチューデントt検定。n=3の生物学的複製物、データは4回の独立した実験を代表。d:主細胞を示す密なチモーゲン果粒を含有するhFGO細胞の透過電子顕微鏡写真。e:MEK阻害因子(PD03)の有無の下でのhAGOと比較したhFGOのペプシノーゲンタンパク質含有量。**:hAGOと比較してp<0.0001、両側スチューデントt検定、hAGO、対照hFGO及びhFGO(PD03非含有)においてそれぞれ、n=8、12、及び11の生物学的複製物。目盛尺:200μm(a)、25μm(b)、及び10μm(d)。エラーバーは、平均値の標準誤差を表す。
hFGOにおける機能的傍細胞の分化を駆動する経路の識別。a:傍細胞遺伝子ATP4、ATP4B、及びGIFの発現は、ベースラインで卵胞と比較して、hFGOの10~100倍の増加を呈したが、PD03/BMP4の2日間のパルスへhFGOを曝露することによって劇的に亢進した。**:hAGOと比較してp<0.05、#:対照hFGOと比較してp<0.05、両側スチューデントt検定、n=4の生物学的複製物、データは15回の独立した実験を代表。b:PD03/BMP4を用いた処理後のATP4B発現傍細胞の刺激された分化。c:hFGO由来の傍細胞は、インビボでの成熟マウス胃底部上皮において認められる傍細胞と類似していた。d:傍細胞を想起させる小管構造を有するhFGO細胞の透過型電子顕微鏡写真。e:ヒト胃底部腺の上皮及びhFGO上皮を、表面上皮におけるMUC5AC発現細胞、及び腺性単位におけるATP4B発現傍細胞へと組織化した。f;SNARF-5Fの管腔注射によるヒスタミンに応じたオルガノイドにおける管腔pHの分析。hFGOにおける管腔pHは迅速に低下したのに対し、hAGOは応答を呈しなかった。オルガノイドをファモチジンまたはオメプラゾールのいずれかで前処理することによって、酸性化を遮断した。hFGO(ヒスタミン)、hFGO(ヒスタミン及びファモチジン)、hFGO(ヒスタミン及びオメプラゾール)、及びhAGO(ヒスタミン)のそれぞれにおけるn=9、9、7、及び4の生物学的複製物、データは3回の独立した実験を代表。g:単離したマウス胃腺及びhFGOにおける小管型パターンでのヒスタミン誘導性アクリジンオレンジ(AO)色素の60分後の蓄積。目盛尺:100μm(b)、10μm(c)、10μm(d)、100μm(e;ヒト胃底部)、20μm(e;hFGO)、及び10μm(g)。エラーバーは、平均値の標準誤差を表す。
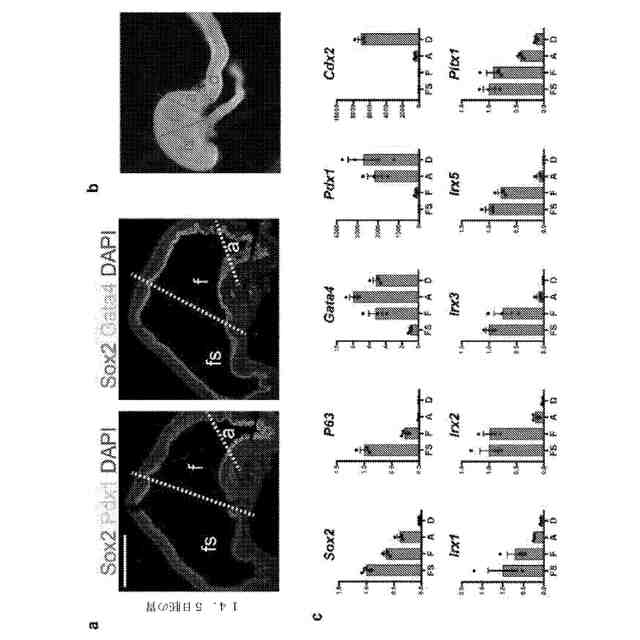
インビボで発達中の胃において規定中の分子ドメイン。a:マウス胚の胃(14.5日胚)におけるSox2、Pdx1、及びGata4の分析は、胃底部(f)がSox2+Gata4+Pdx1-であるのに対し、前庭(a)がSox2+Gata4+Pdx1+であることを示した。前胃(fs)は、Sox2を発現したが、Gata4もPdx1も発現しなかった。b:定量的PCRによって分析した14.5日胚のマウス胃の摘出した領域を示す明視野立体顕微鏡写真。fs:前胃、f:胃底部、a;前庭、d:十二指腸、c:bにおける摘出された領域を、既知の局所的に発現したマーカー(Sox2、P63、Gata4、Pdx1、及びCdx2)についての定量的PCRによって分析して、微量摘出の正確性を確証した。摘出した14.5日胚の胃の領域の定量的PCR分析は、推定上の胃底部マーカーIrx1、Irx2、Irx3、Irx5、及びPitx1は、前庭と比較して胃底部において豊富であることを示した。1箇所の摘出領域当たりのn=4の生物学的複製物。目盛尺:500μm。エラーバーは、標準偏差を表す。
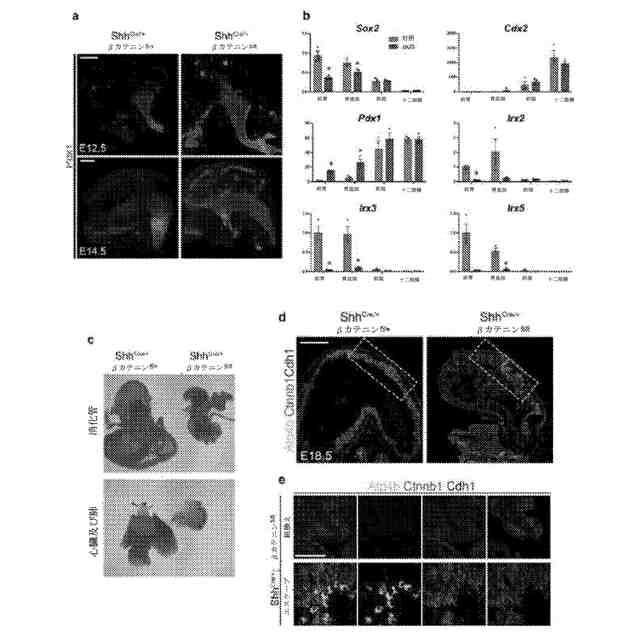
βカテニンcKO胚の分析。a:12.4日胚及び14.5日胚までに、異所性Pdx1発現をcKO胚の背側胃上皮のいたるところ及び最も近位の胃上皮で観察した。b:14.5日胚のcKO前腸の摘出した領域の定量的PCR分析(図6のb)は、胃底部及び前胃部におけるPdx1の有意な上方調節を示した。逆に、Irx2、Irx3、及びIrx5は、これらの近位の領域において顕著に減少した。*:p<0.05、両側スチューデントt検定、各遺伝子型について摘出した領域につきn=3の生物学的複製物。c:18.5日胚から摘出した内臓の立体顕微鏡写真は、cKO胚が既に報告した通り肺の非形成を呈することを実証した。消化管、特に胃は、大きさが劇的に減少した。d:18.5日胚での免疫蛍光染色は、Ctnnb1のモザイク欠失パターンを明らかにした。箱で囲まれた領域を図1に示す。e:18.5日胚のcKO胃において、Ctnnb1染色を欠失する組換え腺は、傍細胞を含有していなかったのに対し、頑強な傍細胞の分化は、Ctnnb1陽性腺において観察された。目盛尺:200μm(a)、500μm(d)、及び50μm(e)。エラーバーは、標準偏差を表す。
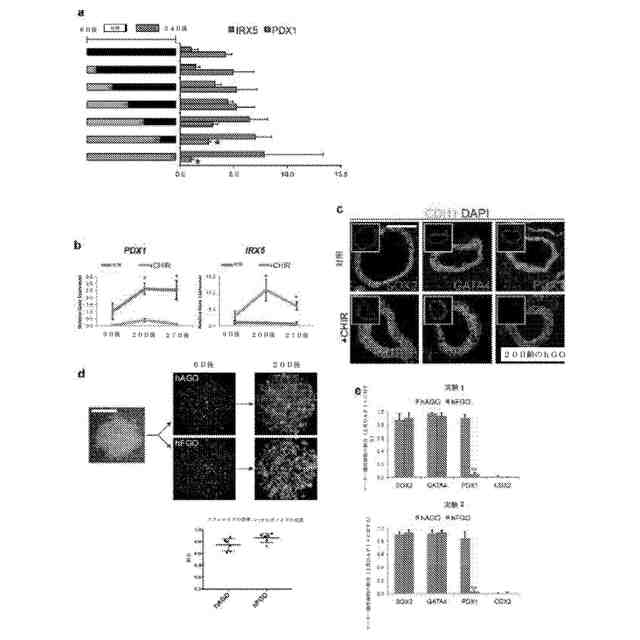
hGOにおける胃底部の運命の適切な誘導及びプロトコルの効率。a:発明者は、CHIR処理が胃底部の識別を確立するのにどれだけ長く必要であるかを研究した。34日後までの対照成長培地中でのオルガノイドの簡潔なCHIR処理(6~9日間)及びその後の成長は結果的に、前庭マーカーPDX1を発現する胃底部オルガノイドを生じ、短期間のCHIR処理が胃底部の運命を安定させないことを示唆した。次に、発明者は、CHIRへのより長い曝露が胃底部の運命を保持するのに必要であるかどうかを検査し、少なくとも29日後までの連続的な処理のみが前庭マーカーPDX1の少量の発現を維持することができることを発見した。*:対照前庭hGOと比較してp<0.05、両側スチューデントt検定。n=3の生物学的複製物、データは2回の独立した実験を代表。b、c:プロトコルの経過にわたって、PDX1は、CHIR処理したオルガノイドにおいて低いままであったのに対し、IRX5発現は、持続的に上昇した。*:p<0.05、両側スチューデントt検定、時点あたりn=3の生物学的複製物。d:6日胚の後部前腸スフェロイドから初期胃オルガノイド(20日胚)への転換は、hAGOプロトコル及びhFGOプロトコルの両方において80%超ほど効率的である。e:20日胚で、hFGO上皮は、約90%のGATA4+/SOX2+/PDX1-であるのに対し、hAGO上皮は、約90%のGATA4+/SOX2+/PDX1+である。**:p<0.001、両側スチューデントt検定、実験あたりn=4の生物学的複製物であって、2回の独立した実験が示されている。目盛尺:100μm(c)及び200μm(d)。
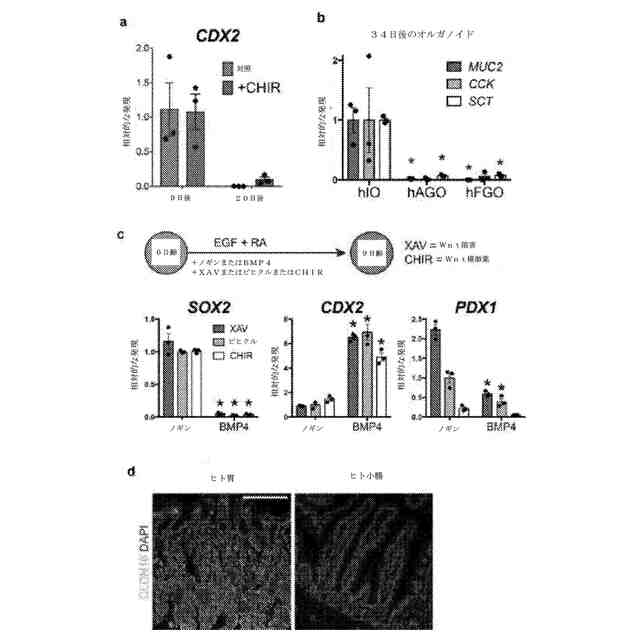
前腸前駆体から小腸の運命を誘導するためのWnt/βカテニン活性化のBMP依存性。a:小腸特異的転写因子CDX2は、9日後も20日後もCHIR処理したhGOにおいて有意に誘導されなかった。b:ヒト小腸オルガノイド(hIO)と比較した場合、胃底部hGOも前庭hGOも、MUC2、CCK、及びSCTを含む、小腸細胞タイプと関連した遺伝子を発現しなかった。*:hIOと比較してp<0.05、両側スチューデントt検定。n=3の生物学的複製物。c:前後の運命は、WNT及びBMPの活性によって協調して制御されている。BMP阻害因子ノギンの存在下で、Wnt/βカテニン経路活性にもかかわらず、オルガノイドはすべて、前腸を維持した(SOX2+)が、BMP4の存在下では、オルガノイドはすべて、後側化した(CDX2+)。BMP阻害状態におけるWnt(CHIR)の活性化は結果的に、胃底部パターンを生じた(SOX2+、PDX1-、CDX2-)のに対し、WNT(CHIR)の活性化及びBMP4の添加は結果的に、小腸の運命を生じた(CDX2+)。*:類似のノギン処理条件と比較してp<0.05、両側スチューデントt検定。n=3の生物学的複製物。d:ヒト組織の免疫蛍光染色は、CLDN18が小腸では認められない胃特異的上皮マーカーであることを明らかにした。目盛尺:200μm。エラーバーは、平均値の標準誤差を表す。
hFGOは、関連する間充織層によって支持される組織化した腺を含有する。a:透過電子顕微鏡写真は、hFGO腺が、狭小尖端膜とともに組織化した構造を呈することを実証した。b:hFGO及びhAGOの両方は、支持層であるFOXF1+/VIM+未分化線維芽細胞を含有していた。目盛尺:5μm(a)及び100μm(b)。
ヒト胃オルガノイドにおける局所特異的細胞分化。a:前庭及び胃底部hGOは、粘膜細胞マーカーMUC5AC及びMUC6のかなりの発現を呈した。b:透過電子顕微鏡写真において示されるように、hFGOは、分化した主細胞への前駆体である粘膜頸細胞と一致した果粒パターンを呈する多量の細胞を含有していた。c:NEUROG3欠乏性hESC株に由来するhGOにおけるNEUROG3の外来性発現は、SYP陽性内分泌細胞の頑強な分化を誘導した。hAGO及びhFGOの両方が、GHRL発現内分泌細胞及びSST発現内分泌細胞を形成したが、GAST+ G細胞の特異性は、hAGOにおいてのみ観察された。d:hGO及びヒト胃生検組織における細胞系譜マーカーの発現比較。定量的PCR分析は、hGOがいくつかの系譜マーカー(MUC5AC、ATP4B)のかなりの発現レベルを呈する一方で、他の遺伝子が、完全に分化した成体ヒト胃において認められるよりも非常に低レベルで発現する(ATP4A、PGA5、及びPGC)ことを実証した。目盛尺:5μm(b)及び100μm(c)。エラーバーは、標準偏差(a)及び平均値の標準誤差(b)を表す。
マウス主細胞発生の分析。a:後期胚期と同じくらい早く機能的マーカー(Atp4b)を発現する傍細胞とは異なり、主細胞遺伝子産物は、発生のかなり後期まで検出することはできなかった。胚(18.5日胚)及び若齢個体(P12)の胃において、Gif及びPgcは、まだ発現しておらず、主細胞が、胃上皮における他の系譜よりも発生上非常に遅く成熟することを示した。b:Pgcがないにもかかわらず、P12マウスの胃は、主細胞特異的マーカーである核MiST1を発現する多量の腺細胞を含有していた。したがって、主細胞は、実際により早期に特異化するが、終末分化マーカーの頑強な発現を発生させるために数週間かかった。目盛尺:100μm(a)及び200μm(b)。
胃底部hGOにおける傍細胞の分化を促進する経路のためのスクリーニング。a:傍細胞の分化を誘導することのできる成長因子/小分子について検査するために、示されたアゴニストまたはアンタゴニストへhFGOを2日間(30~32)曝露した後、34日後に分析した。異なる経路のスクリーニング実験において、PD03を用いたMEK阻害のみが、ATP4A/Bの発現を頑強に誘導することが発見された。b:培地からのEGFの減少または除去は、MEK阻害の効果を再現するのに十分ではなかった。c:傍細胞発生を誘導するPD03/BMP4の能力は、前庭hGOがPD03/BMP4に応じて胃底部マーカーを発現しなかったので、胃底部hGOに限定されていた。d:PD03/BMP4への曝露は、胃底部hGOにおけるATP4A及びATP4Bの発現を迅速に高めた。e:PD03/BMP4を用いた傍細胞発生の誘導は、主細胞(PGA5及びPGC)及び内分泌細胞(CHGA)の分化に有意に影響しなかった。f:hFGO分化プロトコルの各期における操作は、いかなる単一のステップの除去もATP4A/B発現の喪失をもたらしたので、頑強な傍細胞分化に必要であった。エラーバーは、標準偏差(a~c)及び平均値の標準誤差(d~f)を表す。
胃オルガノイドにおける生でのインビトロpHモニタリング。a:色素SNAFR5Fは、5~8のpH範囲にわたって応答性を呈し、このことは、傍細胞仲介性酸分泌に応じた生理学的変化を検出するのに十分適していた。b:ヒスタミン添加の前(黒丸)及び60分後(白丸)に記録した培地及び管腔のpH測定。前庭hGOは、ヒスタミンに対して応答しなかったのに対し、胃底部hGOは、ヒスタミンに応答して管腔pHを低下させた。ファモチジンまたはオメプラゾールのいずれかを用いたオルガノイドの前処理によって、酸性化を阻害した。さらに、オメプラゾールは、ヒスタミン曝露の前に胃底部オルガノイドのpHを上昇させるのに十分であり、このことは、胃底部オルガノイドにおけるベースラインの酸分泌を示唆していた。培地のpHは、いかなるオルガノイドも変化させなかった。***:ヒスタミン前と比較してp<0.001、$$$:ヒスタミンなしでの管腔pHと比較してp<0.001、###:ヒスタミンありでの管腔pHと比較してp<0.001、両側スチューデントt検定。c:hFGOは、腺の管腔を裏打ちする細胞のほぼすべてにおいてアクリジンオレンジ(AO)が蓄積した傍細胞の密な腺を含有していた。d:AO蓄積は、hFGOの傍細胞における小管型パターンで観察された。目盛尺:10μm。エラーバーは、標準偏差を表す。
ヒト胃オルガノイドの連続継代。a:hGOにおける胃幹細胞の存在を判定するための実験の概略図。b:EGFのみを含有する培地中で断片を生育させたとき、新たなオルガノイドを形成するよう生育も増殖もしなかった。しかしながら、培地へのCHIR及びFGF10の添加は、新たに形成されたオルガノイドへの個々の断片の生育を支持するのに十分であった。c:2回の継代後、hFGOは、PGC、MUC6、MUC5AC、及びGHRLを含む、胃表現型と一致する遺伝子をなおも発現した。胃の同一性を維持しながら連続継代を受けるこの能力は、hFGOが、成体胃幹細胞と類似の特性を有する細胞を含有するという結論を支持している。d:継代したhFGOは、いくつかの分化した胃細胞タイプと関係するマーカーを発現したが、ATP4Bなどの傍細胞と関係する遺伝子を発現しなかった。さらに、傍細胞の分化は、継代前ではMEK阻害を通じて誘導することができたのに、MEK阻害を通じて誘導することはできなかった。エラーバーは、標準偏差を表す。
【発明を実施するための形態】
【0007】
別段の記載がない限り、用語は、当業者による従来の使用により理解されるものとする。
【0008】
本明細書で使用する場合、「胃底部組織」という用語は、酸産生傍細胞及びプロテアーゼ産生主細胞を含むがこれらに限定されない胃底部細胞タイプを含有する体において認められる胃上皮の胃底部タイプを意味する。
【0009】
本明細書で使用する場合、「胚体内胚葉(DE)細胞」という用語は、原腸形成の過程によって生じる3つの主要な胚葉のうちの1つを意味する。
【0010】
本明細書で使用する場合、「wntシグナル伝達経路」という用語は、wnt/ベータカテニン経路を意味しており、ベータカテニンタンパク質を通じて作用する、Wntリガンドと縮れた(frizzled)細胞表面の受容体とによって仲介されるシグナル伝達経路である。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
マグネデザイン株式会社
磁気顕微鏡
14日前
個人
セルロース性物質の製造方法
1か月前
株式会社ゴーフォトン
PCR方法
22日前
松谷化学工業株式会社
澱粉分解物の製造方法
2か月前
SMC株式会社
気体供給装置
1か月前
株式会社GSP研究所
miRNA検出方法
2か月前
東ソー株式会社
ポリマーコートビーズの製造方法
2か月前
テルモ株式会社
液体除去器具
4日前
鹿島建設株式会社
褐藻の冷凍保存方法
18日前
サッポロビール株式会社
アルコール飲料
18日前
サッポロビール株式会社
アルコール飲料
1か月前
サッポロビール株式会社
アルコール飲料
1か月前
株式会社テクノーブル
乳酸菌及び皮膚外用剤
18日前
アサヒビール株式会社
容器詰麦芽発酵飲料
1か月前
株式会社SUPER BLOOM
スピリッツ
18日前
テルモ株式会社
移植用デバイス
4日前
新東工業株式会社
培養システム
26日前
新東工業株式会社
培養システム
26日前
株式会社エムスタイル
微生物群の賦活化方法
2か月前
学校法人君が淵学園
核酸の部位特異的アシル修飾剤
1か月前
テルモ株式会社
積層体および移植片の輸送方法
4日前
株式会社渡辺オイスター研究所
脂肪細胞の成長抑制剤
2か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
1か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
1か月前
株式会社ユーグレナ
観察用遠隔制御装置
4日前
サッポロビール株式会社
茶風味アルコール飲料
1か月前
ヤマサ醤油株式会社
アルギニン非分解性乳酸菌株の取得方法
1か月前
株式会社アステック
培養装置及び培養方法
8日前
株式会社関電工
藻類培養装置および藻類培養方法
8日前
学校法人近畿大学
好気性微生物の培養方法および培養器
1か月前
国立大学法人愛媛大学
ASC欠損非ヒト哺乳動物
4日前
TOA株式会社
化粧品の保存効力試験方法
4日前
サムコ株式会社
ポアデバイス
1か月前
花王株式会社
乳酸応答性酵素電極
4日前
宝酒造株式会社
蒸留酒類、及び蒸留酒類の製造方法
4日前
学校法人帝京大学
油脂を過剰蓄積する油脂生産酵母変異株
18日前
続きを見る
他の特許を見る

 特許ウォッチ
特許ウォッチ