TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025016450
公報種別
公開特許公報(A)
公開日
2025-02-04
出願番号
2024168411,2021519861
出願日
2024-09-27,2019-10-11
発明の名称
人工トランス活性化因子による選択
出願人
オスペダーレ サン ラファエレ エス.アール.エル
,
フォンダツィオーネ テレトン イーティーエス
代理人
弁理士法人平木国際特許事務所
主分類
C12N
5/10 20060101AFI20250128BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】遺伝子編集及びin vivo投与の前の編集された細胞の濃縮を達成する方法を提供する。
【解決手段】ゲノム編集された細胞を選択する方法及び/又は細胞の集団におけるゲノム編集された細胞の濃縮のための方法であって、(a)細胞又は細胞の集団中に、少なくとも1つの第1の成分、少なくとも1つの第2の成分及び少なくとも1つの第3の成分を導入すること、並びに(b)セレクターをコードするヌクレオチド配列を一過性に発現するか、又は一過性に上方制御するゲノム編集された細胞を選択することを含む、方法。
【選択図】なし
特許請求の範囲
【請求項1】
ゲノム編集された細胞を選択する方法及び/又は細胞の集団におけるゲノム編集された細胞の濃縮のための方法であって、
(a)細胞又は細胞の集団中に、少なくとも1つの第1の成分、少なくとも1つの第2の成分及び少なくとも1つの第3の成分を導入すること、並びに
(b)セレクターをコードするヌクレオチド配列を一過性に発現するか、又は一過性に上方制御するゲノム編集された細胞を選択すること
を含み、
第1の成分は、セレクターをコードするヌクレオチド配列及び目的のヌクレオチド配列(NOI)及び任意選択で、調節エレメントに作動可能に連結した最小プロモーターを含むドナーレポーターカセットであり、
第2の成分は、操作された転写トランス活性化因子(ETT)ポリペプチド又はETTポリペプチドをコードするヌクレオチド配列であり、ETTポリペプチドは、DNA結合性ドメイン(DBD)及び少なくとも1つの転写活性化因子(TA)ドメインを含み、
第3の成分は、ゲノム標的ヌクレアーゼ及び任意選択で、少なくとも1つの標的とされるゲノム配列を含むガイドRNA(gRNA)を含むヌクレアーゼ系であり、
ETTポリペプチドは、細胞若しくは細胞の集団中に一過性に存在するか、又はETTポリペプチドをコードするヌクレオチド配列は、細胞若しくは細胞の集団において一過性に発現され、及び
細胞又は細胞の集団におけるヌクレアーゼ系の存在が、セレクターをコードするヌクレオチド配列及びNOI及び任意選択で、調節エレメントに作動可能に連結した最小プロモーターの、標的遺伝子座への挿入を可能にし、且つ、任意選択で、モジュレーターが細胞若しくは細胞の集団中に存在する場合に、又は任意選択で、モジュレーターが細胞若しくは細胞の集団中に存在しない場合に、ETTポリペプチドの一過性存在又はETTポリペプチドをコードするヌクレオチド配列の一過性発現が、挿入されたセレクターをコードするヌクレオチド配列の一過性発現又は一過性上方制御を可能にする、
前記方法。
続きを表示(約 2,400 文字)
【請求項2】
ドナーレポーターカセットが順次、
(i)標的遺伝子座と相同なヌクレオチド配列を含む左相同性アーム(HA)、
(ii)最小プロモーターに作動可能に連結したセレクターをコードするヌクレオチド配列、
(iii)プロモーターに作動可能に連結したNOI、及び
(iv)標的遺伝子座と相同なヌクレオチド配列を含む右相同性アーム(HA)
を含み、
セレクターをコードするヌクレオチド配列が標的遺伝子座中に挿入される場合に、第2の成分のETTポリペプチド又は第2の成分によって発現されたETTポリペプチドが、最小プロモーターを活性化する、
請求項1記載の方法。
【請求項3】
ドナーレポーターカセットが順次、
(i)標的遺伝子座と相同なヌクレオチド配列を含む左相同性アーム(HA)、
(ii)任意選択で、スプライシングアクセプター部位(SA)、
(iii)NOI、
(iv)最小プロモーターに作動可能に連結したセレクターをコードするヌクレオチド配列、及び
(v)標的遺伝子座と相同なヌクレオチド配列を含む右相同性アーム(HA)
を含み、
セレクターをコードするヌクレオチド配列が標的遺伝子座中に挿入される場合に、第2の成分のETTポリペプチド又は第2の成分によって発現されたETTポリペプチドが、最小プロモーターを活性化する、
請求項1記載の方法。
【請求項4】
ドナーレポーターカセットが順次、
(i)標的遺伝子座と相同なヌクレオチド配列を含む左相同性アーム(HA)、
(ii)任意選択で、スプライシングアクセプター部位(SA)、
(iii)NOI、
(iv)任意選択で、2A自己切断ペプチド(2A)又は配列内リボソーム進入部位(IRES)エレメントをコードするヌクレオチド配列、
(v)セレクターをコードするヌクレオチド配列、ここで、任意選択で、セレクターをコードするヌクレオチド配列は最小プロモーターに作動可能に連結されている、及び
(vi)標的遺伝子座と相同なヌクレオチド配列を含む右相同性アーム(HA)
を含み、
第2の成分のETTポリペプチド又は第2の成分によって発現されたETTポリペプチドが、標的遺伝子座において内因性プロモーターを活性化する、
請求項1記載の方法。
【請求項5】
ドナーレポーターカセットが順次、
(i)標的遺伝子座と相同なヌクレオチド配列を含む左相同性アーム(HA)、
(ii)NOI、ここで、任意選択で、NOIはプロモーターに作動可能に連結されている、
(iii)最小プロモーターに作動可能に連結された、セレクターをコードするヌクレオチド配列であって、最小プロモーターが調節エレメントに作動可能に連結されている、セレクターをコードするヌクレオチド配列、及び
(iv)標的遺伝子座と相同なヌクレオチド配列を含む右相同性アーム(HA)
を含み、
セレクターをコードするヌクレオチド配列が、標的遺伝子座中に挿入される場合に、及びモジュレーターが細胞又は細胞の集団に存在する場合に、第2の成分のETTポリペプチド又は第2の成分によって発現されたETTポリペプチドが、調節エレメントに結合し、且つ、最小プロモーターを活性化するか、又は
セレクターをコードするヌクレオチド配列が、標的遺伝子座中に挿入される場合に、及びモジュレーターが細胞又は細胞の集団に存在しない場合に、第2の成分のETTポリペプチド又は第2の成分によって発現されたETTポリペプチドが、調節エレメントに結合し、且つ、最小プロモーターを活性化する、
請求項1記載の方法。
【請求項6】
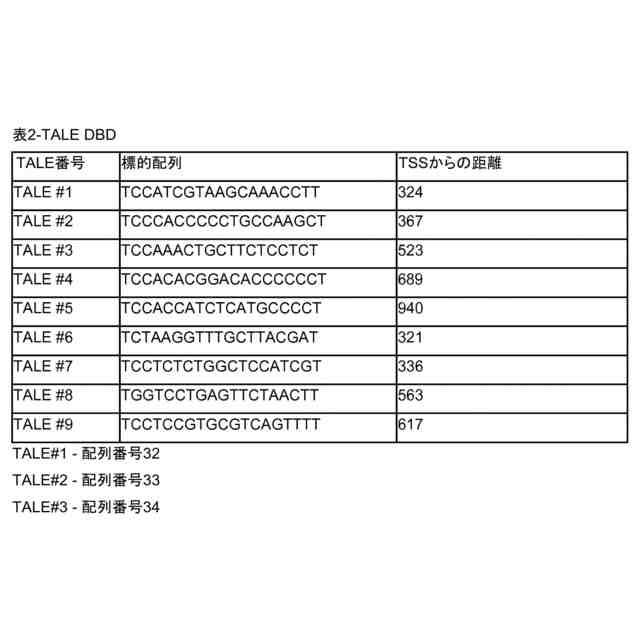
DBDが、転写活性化因子様エフェクター(TALE)DBD、亜鉛フィンガー、触媒的に不活性のCpf1又は触媒的に不活性のCas(dCas)であり、且つ、TAドメインが、VP16、VP64、VP128、VP160、VPR、p65、Rta、HSF1、SAM及びSunTagから成る群から選択される、請求項1~4のいずれか1項記載の方法。
【請求項7】
DBDが、TetR又はリバースTetR(rTetR)であり、且つ、TAドメインが、VP16、VP64、VP128、VP160、VPR、p65、Rta、HSF1、SAM及びSunTagから成る群から選択される、請求項5記載の方法。
【請求項8】
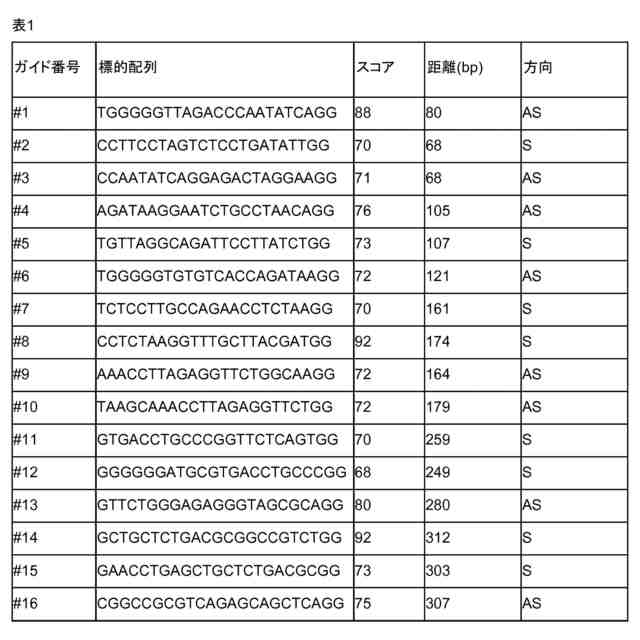
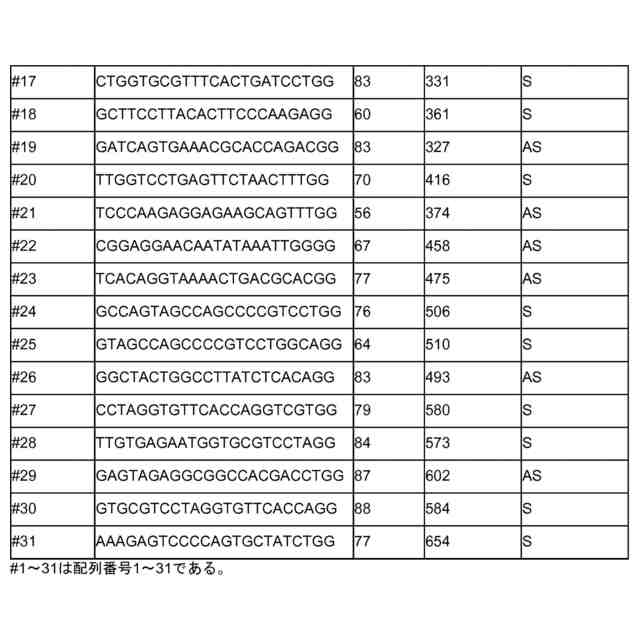
gRNAが、配列番号1~31及びそれと少なくとも75%の同一性を有する配列から成る群から選択されるヌクレオチド配列のうち1つ以上に結合可能である、請求項1~7のいずれか1項記載の方法。
【請求項9】
標的遺伝子座が、セーフハーバーである、請求項1~8のいずれか1項記載の方法。
【請求項10】
標的遺伝子座が、アデノ随伴ウイルス組込み部位1(AAVS1)、レンチウイルスベクターの共通組込み部位(CIS)、IL2RG、gp91phox、HBB、RAG1、CD40LG、TRAC、TRBC、STAT、PRF1、皮膚において発現されるタンパク質(例えば、コラーゲン、ケラチン、ラミニン、デスモコリン、デスモプラチン(desmoplachine)、デスモグレイン、プラコグロビン(placoglobin)、プラコフィリン(placophylline)、インテグリン又はデスモソーム及びヘミデスモソームに関与する他のタンパク質)をコードする遺伝子又は別のセーフハーバーゲノム遺伝子座である、請求項9記載の方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、ゲノム編集された細胞を選択する方法及び/又は細胞の集団におけるゲノム編集された細胞の濃縮のための方法に関する。本発明はまた、方法によって生成されたゲノム編集された細胞の集団、ゲノム編集された細胞の集団を含む医薬組成物、療法(例えば、遺伝子療法、造血幹細胞移植、がん処置及び組織修復)のためのゲノム編集された細胞の集団の使用並びにゲノム編集された細胞の集団を投与するステップを含む、障害、例えば、X連鎖SCID及び皮膚疾患又は網膜疾患の処置又は予防のための方法に関する。
続きを表示(約 2,700 文字)
【背景技術】
【0002】
造血幹細胞(HSC)遺伝子療法は、いくつかの血液疾患及び非血液疾患に冒された患者において実質的な治療利益を提供してきた。それでも、セミランダムに組み込むベクターの使用は、依然として、挿入型突然変異誘発及び非生理学的導入遺伝子発現の危険をもたらす。したがって、HSC遺伝子工学の範囲は、遺伝子置換から、内因性遺伝子の正確な改変のために人工ヌクレアーゼの使用に頼る標的化されたゲノム編集へと広がった。原始造血幹/前駆細胞(HSPC)における遺伝子編集は、何年もとらえどころのないものであった。実際、これらの細胞におけるDNA二本鎖切断(DSB)の誘導は、アポトーシス、分化又は老化を引き起こす場合があり、その修復は、関連タンパク質機構の上方制御及び細胞周期による標的化細胞の進行を必要とする相同組換えDNA修復(HDR)というよりも非相同末端結合(NHEJ)によって進められ得る。
【0003】
ex vivo HSC培養、ヌクレアーゼデザイン及び遺伝子導入技術における顕著な進歩によって、これらの障壁が部分的に克服されることが可能となり、免疫不全NSGマウスにおける長期多系統再構成可能なヒトHSPCにおける標的化ゲノム編集の明確な証拠が提供されている(Genoveseら、2014年)。より最近、遺伝子編集試薬及び手順のさらなる最適化によって、原始HSPCにおいて遺伝子標的化効率が著しく改善され、長期生着HSPCにおいて平均で10~20%の遺伝子マーキングを達成した(De Ravinら、2017年;Deverら、2016年;Wangら、2015年)。遺伝子修正のこれらのレベルは、いくつかの疾患の処置にとって、例えば、X連鎖重症複合免疫不全症(SCID-X1)の場合において安全であり、有効であると予測されており、編集された後代は、編集されていない対応物を上回って選択的利点の利益を有するであろう(Schiroliら、2017年)。
【0004】
しかし、より広い臨床適用にとって適したものになる前にHSC遺伝子編集が直面するいくつかの課題がある。特に、より原始のHSPCにおける遺伝子編集の効率並びに編集されたHSCの収率を改善することは重大である。遺伝子編集プロトコールのさらなる改善は、遺伝子標的化効率を増強し得るが、投与前の修正されたHSPCのin vitro選択は、編集された細胞の純粋な集団を得るための価値ある戦略であろう。編集された細胞の選択は、遺伝子修正のレベルが病理過程の復帰を達成するように十分でない場合には困難である。さらに、修正された遺伝子が成長の利点を提供しない場合がある(例えば、慢性肉芽腫性疾患、サラセミア、RAG1欠損症などにおいて生じる)ので、意図された編集を直接選択することは困難であり得る。さらに、遺伝子編集された細胞は、その関連タンパク質産物の細胞内局在又はタンパク質がHSPCにおいて発現されない場合さえあるために、選択に適していない場合がある。
【0005】
これらのハードルを克服するために、遺伝子編集を、選択マーカー、例えば、FACS選別に適している蛍光タンパク質又は薬物耐性タンパク質の構成的発現と、それに続く選択を媒介するための薬物処置と合わせてもよい。しかし、レポーター遺伝子の構成的発現は免疫原性である、又は長期細胞生存度にとって有害である場合があり、したがって、改変された細胞の喪失につながる可能性がある。最後に、これらのシステムは、インサイツ(in situ)及びオフサイト(off-site)両方の標的化された挿入の選択につながり、したがって、治療及び安全性の結果を危険にさらす。
【0006】
操作された転写トランス活性化因子(ETT)のブレークスルーによって、意図される遺伝子の発現を正確に調節する新しい可能性を開かれた。特に、TALE及びCRISPR/Cas系が、それぞれ、DNA結合性ドメイン(DBD)又は触媒的に不活性のCas9(dCas9)を、単一又は複数の転写活性化因子ドメイン(例えば、VP64及びVPR)と融合することによって設計された(Chavezら、2015年)。
【0007】
Guoら(2017年、Protein Cell 8巻(5号):379~393頁)では、ヒト多能性幹細胞(HPSC)において内因性遺伝子を上方制御する系が論じられている。Guoら(2017年)では、ドキシサイクリン(Dox)誘導型dCas9-VP64-p65-Rta及びTetトランス活性化因子発現カセットの、AAVS1遺伝子座の対立遺伝子中への挿入並びにdCas9-VPRのレベルが、Doxの付加及び撤回によって正確に、可逆的に制御され得ることが開示されている。
【0008】
Deverら(2016年、Nature 539巻(7629号):384~389頁)では、Cas9リボヌクレオタンパク質及び組換え体AAV6相同組換えドナー送達の組合せを使用することによる、ヒト幹細胞(HSC)におけるHBB遺伝子での相同組換えが論じられている。Deverら(2016年)では、tNGFRを含むAAVベクタープラスミドが開示されている。Deverら(2016年)では、HBB標的化HSPCを濃縮するためのマーカーとしてのtNGFR及び抗NGFR磁性ビーズ分離を使用することが論じられている。
【0009】
Bakら(2018年、Nat Protoc 13巻(2号):358~376頁)では、相同組換えによってヒト造血幹細胞を編集するためのCRISPR/Cas9技術及び組換えAAV6相同ドナー送達の使用が論じられている。Bakら(2018年)には、レポーター遺伝子、例えば、GFP又は切断型増殖因子受容体(tNGFR)が開示されている。Bakら(2018年)には、標的化された組込みを有する細胞を濃縮するためのフローサイトメトリーが論じられている。
【0010】
US2015/0191744には、ヌクレアーゼ欠損Cas9エフェクタードメイン融合タンパク質をコードするレンチウイルスベクター及びゲノム標的と相補的である少なくとも1つのsgRNA遺伝子を含むレンチウイルスベクターを使用して幹細胞又は前駆細胞の転写調節を改変する方法が論じられている。US2015/0191744には、Cas9-蛍光タンパク質融合タンパク質が開示されている。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
株式会社ゴーフォトン
PCR方法
7日前
個人
セルロース性物質の製造方法
1か月前
SMC株式会社
気体供給装置
17日前
株式会社GSP研究所
miRNA検出方法
1か月前
鹿島建設株式会社
褐藻の冷凍保存方法
3日前
サッポロビール株式会社
アルコール飲料
1か月前
株式会社テクノーブル
乳酸菌及び皮膚外用剤
3日前
サッポロビール株式会社
アルコール飲料
3日前
サッポロビール株式会社
アルコール飲料
1か月前
アサヒビール株式会社
容器詰麦芽発酵飲料
1か月前
新東工業株式会社
培養システム
11日前
学校法人君が淵学園
核酸の部位特異的アシル修飾剤
1か月前
新東工業株式会社
培養システム
11日前
株式会社エムスタイル
微生物群の賦活化方法
1か月前
株式会社SUPER BLOOM
スピリッツ
3日前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
1か月前
サッポロビール株式会社
茶風味アルコール飲料
19日前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
1か月前
ヤマサ醤油株式会社
アルギニン非分解性乳酸菌株の取得方法
24日前
学校法人近畿大学
好気性微生物の培養方法および培養器
1か月前
サムコ株式会社
ポアデバイス
1か月前
国立大学法人富山大学
T細胞受容体およびその利用
1か月前
株式会社豊田中央研究所
環境核酸モニタリング方法
1か月前
学校法人帝京大学
油脂を過剰蓄積する油脂生産酵母変異株
3日前
雪印メグミルク株式会社
D-セリン濃度低減用組成物
2か月前
三浦工業株式会社
試料液に含まれる微生物の生体の抽出方法
1か月前
学校法人帝京大学
細胞周期同調が可能な油脂生産酵母変異株
3日前
個人
液滴処理装置、液滴処理システム及び液滴処理方法
25日前
学校法人関西医科大学
ポリペプチド
1か月前
個人
細胞処理装置、細胞処理システム及び細胞処理方法
1か月前
グリーンケミカルズ株式会社
フェノール組成物およびポリマー
2か月前
花王株式会社
目的タンパク質の生産方法
1か月前
ニッタ株式会社
凍結細胞解凍装置
2か月前
ヤマト科学株式会社
生体由来物質の濃縮方法
24日前
サントリーホールディングス株式会社
ビールテイスト飲料
1か月前
学校法人東邦大学
自己反応性T細胞の検出方法
2か月前
続きを見る
他の特許を見る



 特許ウォッチ
特許ウォッチ