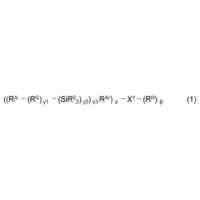TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2024073492
公報種別
公開特許公報(A)
公開日
2024-05-29
出願番号
2024032219,2020195185
出願日
2024-03-04,2016-10-21
発明の名称
セストリン-GATOR2相互作用のモジュレーターおよびその使用
出願人
ナビター ファーマシューティカルズ, インコーポレイテッド
代理人
個人
,
個人
,
個人
,
個人
,
個人
主分類
C07C
229/12 20060101AFI20240522BHJP(有機化学)
要約
【課題】化合物、その組成物、およびそれらを使用する方法を提供すること
【解決手段】本発明は、化合物、その組成物、およびそれらを使用する方法を提供する。本発明は、セストリン-GATOR2相互作用をモジュレートし、これにより間接的に、mTORC1活性を選択的にモジュレートするために有用な化合物および方法に関する。本発明はまた、本発明の化合物を含む薬学的に許容される組成物および様々な障害の処置において前記組成物を使用する方法も提供する。本発明の化合物および薬学的に許容されるその組成物は、mTORC1に関連する、様々な疾患、障害または状態の処置に有用である。このような疾患、障害または状態には、本明細書に記載されているものなどの、糖尿病、てんかん、神経変性、免疫応答、骨格筋成長の抑制および細胞増殖性障害(例えば、がん)が含まれる。
【選択図】なし
特許請求の範囲
【請求項1】
明細書に記載の発明
。
発明の詳細な説明
【技術分野】
【0001】
発明の技術分野
本発明は、セストリン-GATOR2相互作用をモジュレートし、これにより間接的に、mTORC1活性を選択的にモジュレートするために有用な化合物および方法に関する。本発明はまた、本発明の化合物を含む薬学的に許容される組成物および様々な障害の処置において前記組成物を使用する方法も提供する。
続きを表示(約 8,000 文字)
【背景技術】
【0002】
発明の背景
ラパマイシン複合体1(mTORC1)タンパク質キナーゼの機構的標的は、増殖因子、細胞ストレス、ならびに栄養素およびエネルギーレベルなどの、多様な環境的合図を感じるマスター成長調節因子である。mTORC1は、活性化されると、mRNA翻訳および脂質合成などの同化作用を賦活する基質をリン酸化し、オートファジーなどの異化作用を制限する。mTORC1の調節異常は、とりわけ、糖尿病、てんかん、神経変性、免疫応答、骨格筋成長の抑制およびがんを含めた、幅広い範囲の疾患において発生する(Howellら、(2013年)Biochemical Society transactions 41巻、906~912頁;Kimら、(2013年) Molecules and cells 35巻、463~473頁;LaplanteおよびSabatini、(2012年) Cell 149巻、274~293頁)。
【0003】
増殖因子およびエネルギーレベルを含めた多数の上流での入力により、TSC複合体を介して、mTORC1にシグナルが伝達され、このTSC複合体は、mTORC1の必須のアクチベータである小さなGTPアーゼである、Rhebを調節する(Brugarolasら、(2004年)Genes &;Development 18巻、2893~2904頁;Garamiら、(2003年)Molecular Cell 11巻、1457~1466頁;Inokiら、(2003年)Genes &;Development 17巻、1829~1834頁;Longら、(2005年)Current Biology 15巻、702~713頁;Sancakら、(2008年)Science(New York、NY) 320巻、1496~1501頁;Saucedoら、(2003年)Nature cell biology 5巻、566~571頁;Stockerら、(2003年)Nature cell biology 5巻、559~565頁;Teeら、(2002年)Proc Natl Acad Sci USA
99巻、13571~13576頁)。アミノ酸は、TSC-Rheb軸を介してmTORC1にシグナル伝達しないように思われ、その代わりに、それぞれ、RagCまたはRagDに結合しているRagAまたはRagBからなるヘテロ二量体Rag GTPアーゼを介して作用する(Hiroseら、(1998年) Journal of cell science 111巻(第1部)、11~21頁;Kimら、(2008年) Nature cell biology 10巻、935~945頁;Nobukuniら、(2005年) Proc Natl Acad Sci USA 102巻、14238~14243頁;Roccioら、(2005年) Oncogene 25巻、657~664頁;Sancakら、(2008年) Science(New York、NY)320巻、1496~1501頁;Schuermannら、(1995年) The Journal of biological chemistry 270巻、28982~28988頁;Sekiguchiら、(2001年) The Journal of biological chemistry 276巻、7246~7257頁;Smithら、(2005年) The Journal of biological chemistry 280巻、18717~187
27頁)。Rag GTPアーゼは、mTORC1の細胞内局在化を制御しており、アミノ酸は、Rheb GTPアーゼがやはり生存しているリソソーム表面にmTORC1の動員を促進する(Buergerら、(2006年) Biochemical and Biophysical Research Communications 344巻、869~880頁;Dibbleら、(2012年) Molecular cell 47巻、535~546頁;Saitoら、(2005年) Journal of Biochemistry 137巻、423~430頁;Sancakら、(2008年) Science(New Yor
k、NY)320巻、1496~1501頁)。Rag GTPアーゼの上流経路のいくつ
かの正の構成成分が特定されている。Ragulator複合体は、Rag GTPアーゼをリソソーム表面に局在化させて、液胞型ATPアーゼと一緒になって、RagA/B上でのGDPのGTPへの交換を促進する(Bar-Peledら、(2012年) Cell 15
0巻、1196~1208頁;Sancakら、(2010年) Cell 141巻、290~303頁;Zoncuら、(2011年)Science Signaling 334巻、678~683頁)
。別個のFLCN-FNIP複合体が、RagC/D上で作用し、GTPのGDPへの加水分解を刺激する(Tsunら、2013年)。RagA/BがGTPにロードされ、RagC/DがGDPにロードされると、それらのヘテロ二量体が結合して、そのアクチベータであるRheb GTPアーゼに接触することができるようになる、リソソーム表面にmTORC1を動員する。
【0004】
最近の研究により、アミノ酸のセンシング経路の主要な負の調節因子として、GATOR1多タンパク質複合体が特定され、その喪失により、mTORC1シグナル伝達がアミノ酸飢餓に対して完全に無応答となることを引き起こす(Bar-Peledら、(2013年)Science 340巻、1100~1106頁;Panchaudら、(2013年)Science Signaling 6巻、ra42)。GATOR1は、DEPDC5、Nprl2およびNprl3からなり、RagA/Bに対するタンパク質(GAP)を活性化するGTPアーゼである。GATOR2多タンパク質複合体は、5種の公知のサブユニット(WDR24、WDR59、Mios、Sec13およびSeh1L)を有しており、GATOR1の経路およびその上流の、またはGATOR1に並行する正の構成成分であるが、その分子機能は、最近まで未知であった(Bar-Peledら、(2013年)Science 340巻、1100~1106頁)。
【0005】
最近、1つまたは複数のセストリンにGATOR2が結合していることが特定されたことによって、mTORC1経路に関する追加的な情報が明らかにされ、生じたセストリン-GATOR2複合体が、mTORC1の細胞内局在化および活性を調節することが実証された。特に、セストリン-GATOR2複合体が存在すると、mTORC1経路が阻害され、mTORC1のリソソーム膜への転座が阻止されることにより、mTORC1活性が低下する。GATOR2とセストリン、特にセストリン1およびセストリン2との相互作用は、アミノ酸、特にロイシンにより拮抗され、それほど高い程度ではないが、イソロイシン、メチオニンおよびバリンにより拮抗される。ロイシンの存在下では、GATOR2は、セストリン1またはセストリン2と相互作用をすることはなく、mTORC1は、mTORC1が活性な状態となるリソソーム膜に移動することができる。セストリン1およびセストリン2は、ロイシンに直接結合し、それほど高い程度ではないが、イソロイシンおよびメチオニンに直接結合する(Chantranupongら、(2014年) Cell Rep.;
9巻(1号):1~8頁)。セストリン1または2によるロイシンの結合は、GATOR2との相互作用、およびその後のmTORC1の活性化の撹乱に必要とされる。ロイシンに結合することができないセストリン2変異体は、ロイシンの存在をmTORC1にシグナル伝達することができず、セストリン2およびそのホモログが欠損している細胞は、mTORC1をロイシンの非存在に対して無応答にする(Wolfsonら、(2015年) Science pii:ab2674[印刷前のEpub])。
【0006】
セストリンは、十分に特徴付けられていない分子機能の3種の関連タンパク質(セストリン1、2および3)である(Buckbinderら、(1994年)Proc Natl Acad Sci USA 91巻、10640~10644頁;Budanovら、(2002年)Cell 134巻、
451~460頁;Peetersら、(2003年) Human genetics 112巻、573~580頁)。セストリン2は、mTORC1のシグナル伝達を阻害し、TSCの上流のAMPKを活性化して、TSCと相互作用する(BudanovおよびKarin、(2008年) Cell 134巻、451~460頁)ことが提唱されているが、後の研究により、AMPK
の非存在下で、セストリン2によりmTORC1が阻害されることが見出され(Pengら、(2014年) Cell 159巻(1号):122~33頁)、このことは、GATOR2複合体が、セストリン2に応答して、mTORC1をモジュレートする際に果たす重要な役割をさらに強調するものである。
セストリン-GATOR2複合体のモジュレートは、間接的に、mTORC1活性を選択的にモジュレートするための潜在的な治療標的となる。
【先行技術文献】
【非特許文献】
【0007】
Howellら、(2013年)Biochemical Society transactions 41巻、906~912頁
Kimら、(2013年) Molecules and cells 35巻、463~473頁
LaplanteおよびSabatini、(2012年) Cell 149巻、274~293頁
Brugarolasら、(2004年)Genes &;Development 18巻、2893~2904頁
Garamiら、(2003年)Molecular Cell 11巻、1457~1466頁
Inokiら、(2003年)Genes &;Development 17巻、1829~1834頁
Longら、(2005年)Current Biology 15巻、702~713頁
Sancakら、(2008年)Science(New York、NY) 320巻、1496~1501頁
Saucedoら、(2003年)Nature cell biology 5巻、566~571頁
Stockerら、(2003年)Nature cell biology 5巻、559~565頁
Teeら、(2002年)Proc Natl Acad Sci USA 99巻、13571~13576頁
Hiroseら、(1998年) Journal of cell science 111巻(第1部)、11~21頁
Kimら、(2008年) Nature cell biology 10巻、935~945頁
Nobukuniら、(2005年) Proc Natl Acad Sci USA 102巻、14238~14243頁
Roccioら、(2005年) Oncogene 25巻、657~664頁
Sancakら、(2008年) Science(New York、NY)320巻、1496~1501頁
Schuermannら、(1995年) The Journal of biological chemistry 270巻、28982~28988頁
Sekiguchiら、(2001年) The Journal of biological chemistry 276巻、7246~7257頁
Smithら、(2005年) The Journal of biological chemistry 280巻、18717~18727頁
Buergerら、(2006年) Biochemical and Biophysical Research Communications 344巻、869~880頁
Dibbleら、(2012年) Molecular cell 47巻、535~546頁
Saitoら、(2005年) Journal of Biochemistry 137巻、423~430頁
Sancakら、(2008年) Science(New York、NY)320巻、1496~1501頁
Bar-Peledら、(2012年) Cell 150巻、1196~1208頁
Sancakら、(2010年) Cell 141巻、290~303頁
Zoncuら、(2011年)Science Signaling 334巻、678~683頁
Bar-Peledら、(2013年)Science 340巻、1100~1106頁
Panchaudら、(2013年)Science Signaling 6巻、ra42
Chantranupongら、(2014年) Cell Rep.;9巻(1号):1~8頁
Wolfsonら、(2015年) Science pii:ab2674[印刷前のEpub]
Buckbinderら、(1994年)Proc Natl Acad Sci USA 91巻、10640~10644頁
Budanovら、(2002年)Cell 134巻、451~460頁
Peetersら、(2003年) Human genetics 112巻、573~580頁
BudanovおよびKarin、(2008年) Cell 134巻、451~460頁
Pengら、(2014年) Cell 159巻(1号):122~33頁
【発明の概要】
【課題を解決するための手段】
【0008】
発明の要旨
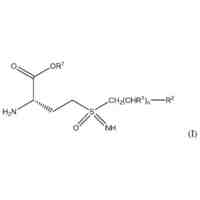
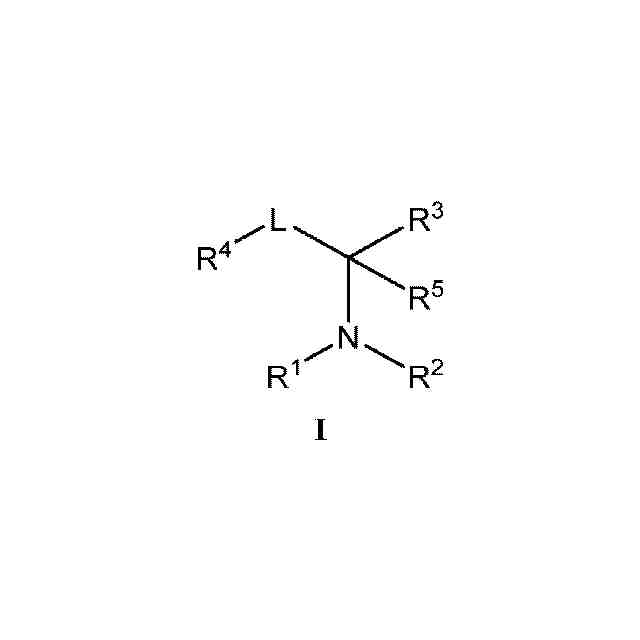
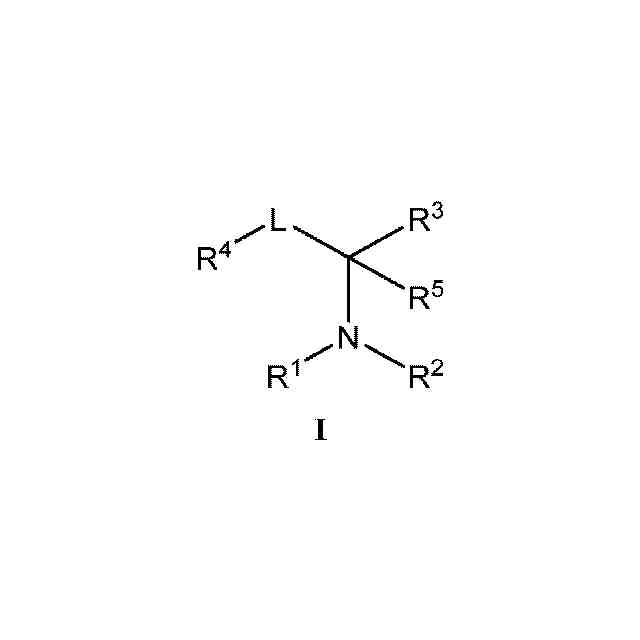
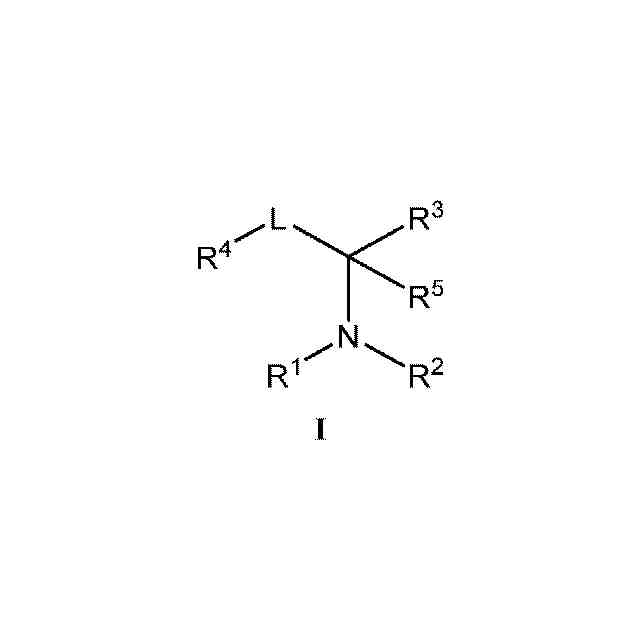
本発明の化合物および薬学的に許容されるその組成物は、セストリン-GATOR2モジュレーターとして有効であることが今や見出された。このような化合物は、一般式I:
TIFF
2024073492000001.tif
36
46
または薬学的に許容されるその塩を有しており、可変基はそれぞれ、本明細書に定義および記載されている通りである。
【0009】
本発明の化合物および薬学的に許容されるその組成物は、mTORC1に関連する、様々な疾患、障害または状態の処置に有用である。このような疾患、障害または状態には、本明細書に記載されているものなどの、糖尿病、てんかん、神経変性、免疫応答、骨格筋成長の抑制および細胞増殖性障害(例えば、がん)が含まれる。
【発明を実施するための形態】
【0010】
ある特定の実施形態の詳細な説明
1.本発明のある特定の実施形態の一般的な説明:
本発明の化合物およびその組成物は、セストリン-GATOR2モジュレーターとして有用である。ある特定の実施形態では、本発明は、式Iの化合物:
TIFF
2024073492000002.tif
40
43
または薬学的に許容されるその塩(式中、
R
1
は、HまたはC
1~6
アルキルであり、
R
2
は、R、-(CH
2
)
n
-フェニル、-C(O)R、-SO
2
Rまたは-C(O)N(R)
2
であり、
nは、0、1または2であり、
Rはそれぞれ独立して、水素、-CN、あるいは飽和もしくは不飽和C
1~6
脂肪族、フェニル、4~7員の飽和もしくは部分不飽和の単環式炭素環式環、1~4個のヘテロ原子を有する5~6員の単環式ヘテロアリール環、または窒素、酸素もしくは硫黄から独立して選択される1~2個のヘテロ原子を有する4~8員の飽和または部分飽和の複素環式環から選択される、任意選択で置換されている基であり、
R
3
は、環A、-C(O)R、-C(O)OR、-C(O)N(R)
2
、-SO
3
H、-SO
2
N(R)
2
、-S(O)R、-S(O)環A、-ORまたは-B(OR)
2
であり、この場合、同一ホウ素上の2つのOR基はそれらの介在原子と一緒になって、ホウ素および2個の酸素に加えて、窒素、酸素もしくは硫黄から独立して選択される0~3個のヘテロ原子を有する、5~8員の飽和もしくは部分不飽和の単環式環を形成するか、またはR
3
とR
4
が一緒になって、窒素、酸素もしくは硫黄から選択される0~1個のヘテロ原子を有する、任意選択で置換されている5~6員環を形成し、
Lは、共有結合、または1~9つのフルオロ基により任意選択で置換されている直鎖もしくは分枝状C
1~6
アルキレン鎖であり、
環Aは、フェニル、または窒素、酸素もしくは硫黄から独立して選択される1~4個のヘテロ原子を有する、任意選択で置換されている5~6員のヘテロアリール環から選択される、任意選択で置換されている環であり、
R
4
は、R、-CF
3
、-OR、-N(R)
2
、-Si(R)
3
もしくは-SRであるか、またはR
3
とR
4
が一緒になって、窒素、酸素もしくは硫黄から選択される0~1個のヘテロ原子を有する、任意選択で置換されている5~6員環を形成し、
R
5
は、HまたはC
1~4
アルキルである)
を提供する。
2.化合物および定義:
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
小野薬品工業株式会社
GCL阻害剤
14日前
築野グループ株式会社
脂肪酸エステル
2か月前
築野グループ株式会社
脂肪酸エステル
2か月前
日本精化株式会社
ホスホコリン誘導体の製造方法
3か月前
ダイキン工業株式会社
シラン化合物
1か月前
ダイキン工業株式会社
シラン化合物
2か月前
東ソー株式会社
免疫グロブリン吸着剤の製造方法
2か月前
田岡化学工業株式会社
多環芳香族化合物の製造方法
20日前
学校法人近畿大学
アミロイド線維検出プローブ
1か月前
株式会社トクヤマ
ベンジルヒドラジン誘導体の製造方法
22日前
三菱ケミカル株式会社
アセトキシアリル化合物の製造方法
3か月前
個人
希土類有機錯体及びその製造方法並び応用
13日前
ヤマナカヒューテック株式会社
反応生成物の製造方法
2か月前
株式会社半導体エネルギー研究所
有機金属錯体、発光デバイス
1か月前
日本特殊陶業株式会社
反応装置
2か月前
東ソー株式会社
アデノ随伴ウイルス結合性タンパク質の精製方法
27日前
住友精化株式会社
アミノシルセスキオキサン
3か月前
グリーンケミカルズ株式会社
4-ヒドロキシ安息香酸組成物
15日前
株式会社半導体エネルギー研究所
有機化合物、および発光デバイス
20日前
東亞合成株式会社
ペプチドフラグメント及びその利用
1か月前
三菱ケミカル株式会社
ポリシロキサン及びその製造方法
2か月前
サンアプロ株式会社
金属錯塩色素
28日前
国立大学法人富山大学
抗がん作用を有する化合物
1か月前
株式会社Kyulux
有機発光素子
15日前
ノーベルファーマ株式会社
ヒスチジン亜鉛2水和物の製造方法
1か月前
ポリプラスチックス株式会社
ホルムアルデヒドの回収方法
2か月前
東ソー株式会社
抗体結合性タンパク質固定化担体を用いた抗体精製方法
3か月前
株式会社日本触媒
アクリル酸の製造方法
2か月前
興和株式会社
新規ニコチンアミド化合物
28日前
ポリプラスチックス株式会社
ホルムアルデヒドの回収方法
2か月前
東洋エンジニアリング株式会社
尿素の製造方法及び製造装置
2か月前
学校法人日本大学
化合物、腫瘍崩壊剤及び医薬組成物
2か月前
積水メディカル株式会社
アルギニン誘導体
今日
イーライ リリー アンド カンパニー
KRAS阻害剤
2か月前
国立大学法人北海道大学
酵素を用いたペプチドライゲーション
1か月前
株式会社ナリス化粧品
バラ花弁由来グルコシルセラミド
1か月前
続きを見る
他の特許を見る




 特許ウォッチ
特許ウォッチ