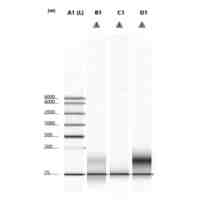
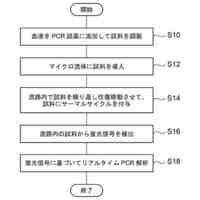
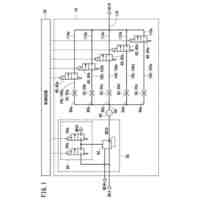
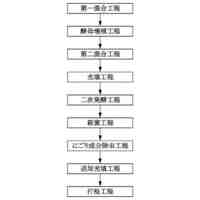
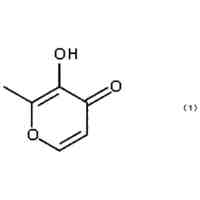
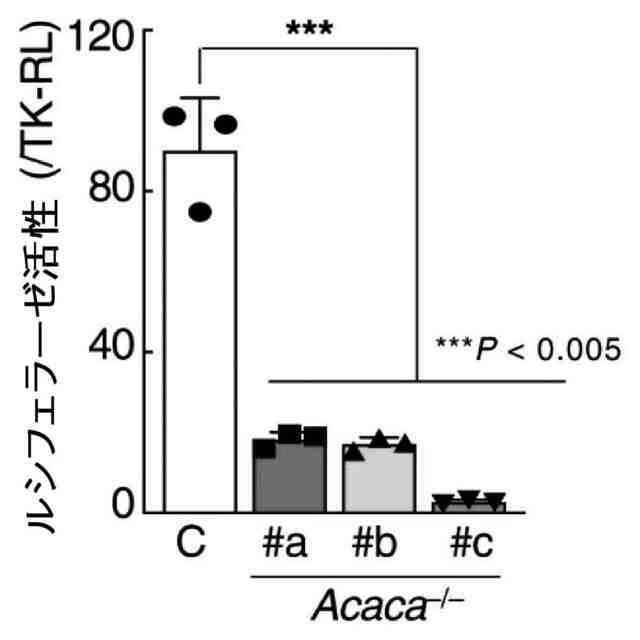
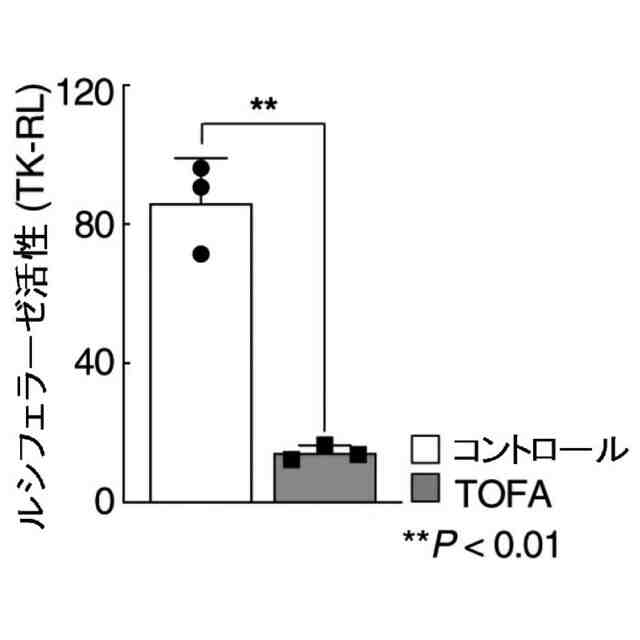
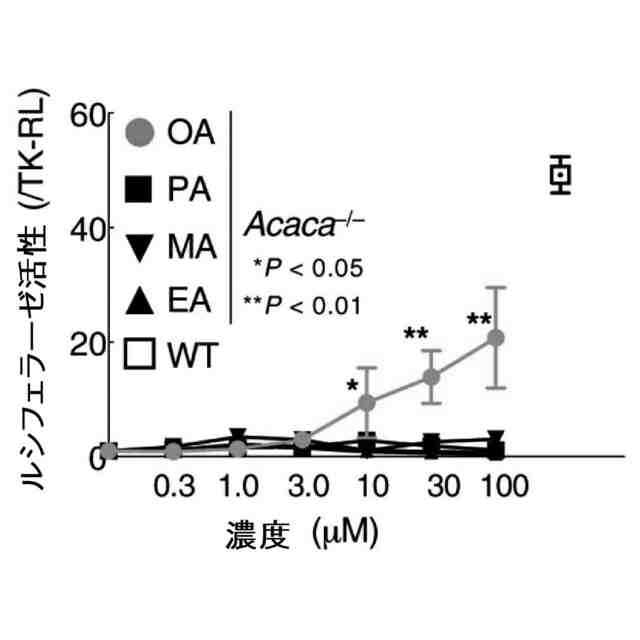
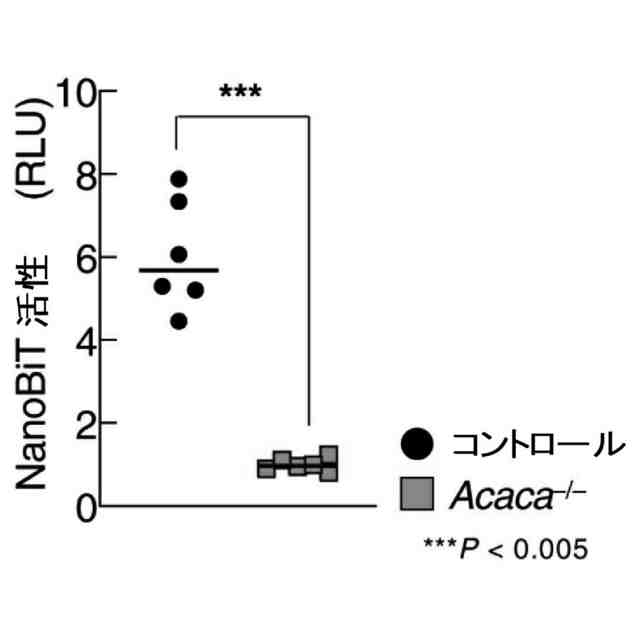
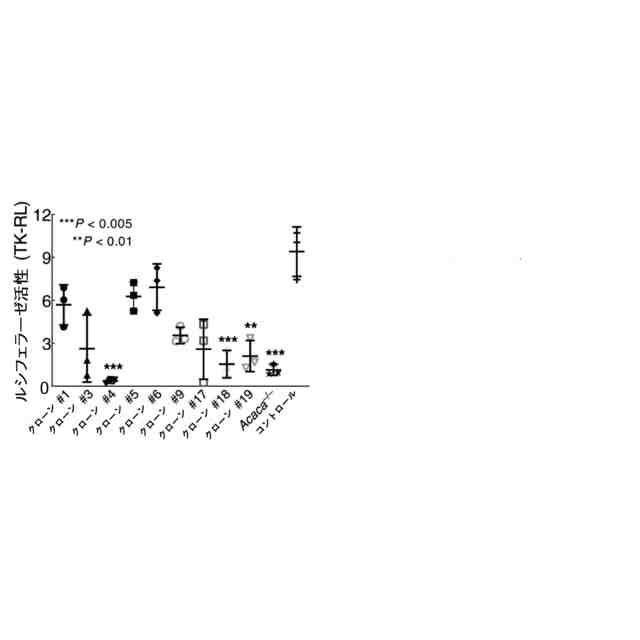
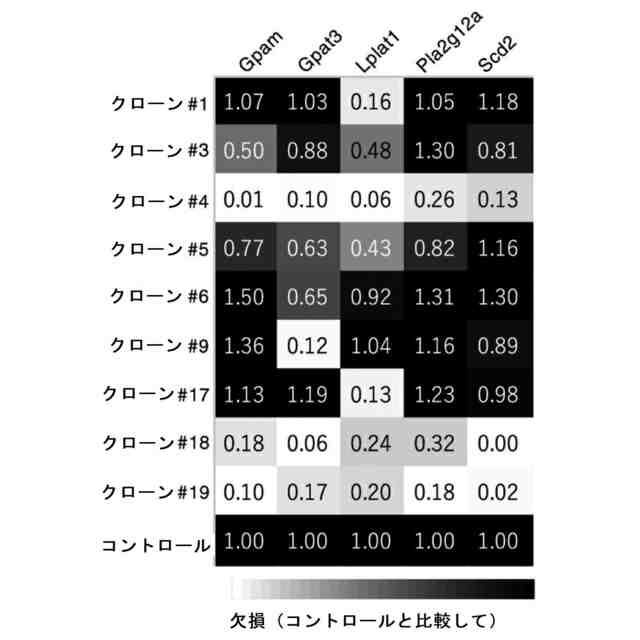
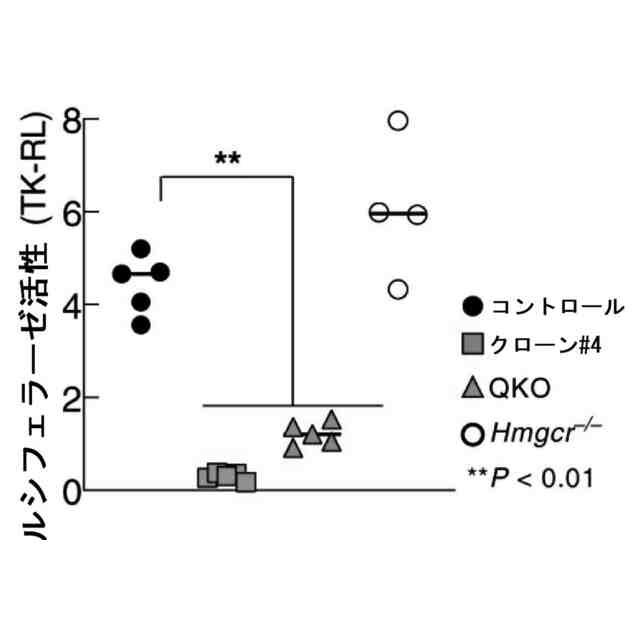
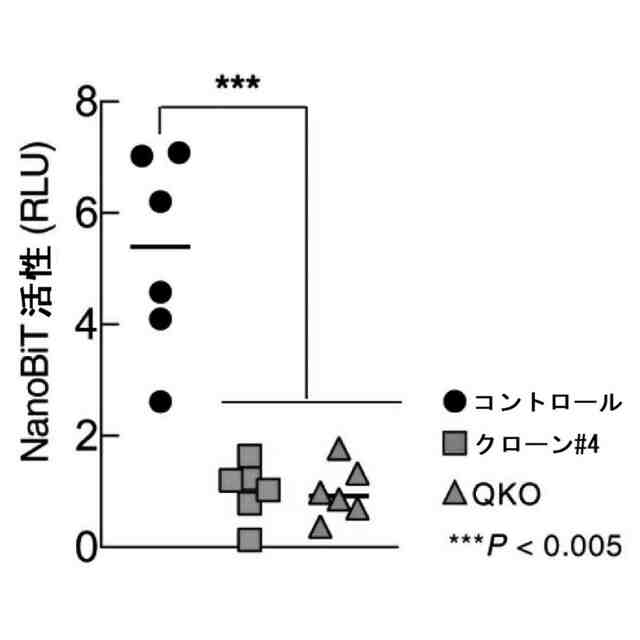
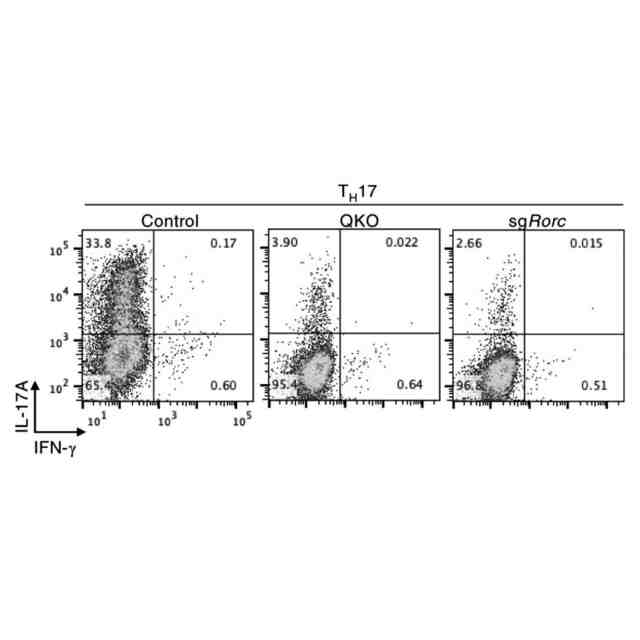
特許請求の範囲【請求項1】 ナイーブCD4陽性T細胞を、1-オレオイル-リゾホスファチジルエタノールアミン、IL-6及びTGF-βを含有する培地中で培養することにより、ナイーブCD4陽性T細胞のTh17細胞への分化を誘導することを含む、Th17細胞の製造方法。 続きを表示(約 1,100 文字)【請求項2】 ナイーブ表現型CD4陽性T細胞が、Pla2g12a、Lplat1、Gpam、Gpat3、Scd2及びACC1からなる群から選択される少なくとも1つの脂質代謝酵素遺伝子の欠損、変異又は発現不全を有するか、Th17細胞へ分化する過程のT細胞においてPla2g12a、Lplat1、Gpam、Gpat3、Scd2及びACC1からなる群から選択される少なくとも1つの脂質代謝酵素遺伝子の発現が抑制される、請求項1記載の製造方法。 【請求項3】 1-オレオイル-リゾホスファチジルエタノールアミンを含有する、Th17細胞分化促進剤。 【請求項4】 1-オレオイル-リゾホスファチジルエタノールアミン及びTh17細胞を含む、細胞調製物。 【請求項5】 1-オレオイル-リゾホスファチジルエタノールアミン及びRORγtを含む、組み合わせ物。 【請求項6】 1-オレオイル-リゾホスファチジルエタノールアミンとRORγtとの相互作用を阻害する化合物を選択することを含む、Th17細胞の活性化に起因する疾患の予防又は治療剤の候補物質のスクリーニング方法。 【請求項7】 Pla2g12a、Lplat1、Gpam、Gpat3、及びScd2からなる群から選択されるいずれかの脂質代謝酵素の活性を阻害する化合物を選択することを含む、Th17細胞の活性化に起因する疾患の予防又は治療剤の候補物質のスクリーニング方法。 【請求項8】 脂質代謝酵素がPla2g12aであり、 被検化合物の存在下で、1-オレオイルホスファチジルエタノールアミンをPla2g12aとともにインキュベートし、生成された1-オレオイル-リゾホスファチジルエタノールアミンを定量し、該生成量が被検化合物の非存在下での生成量を下回る場合に、該被検化合物をTh17細胞の活性化に起因する疾患の予防又は治療剤の候補物質として選択する、請求項7記載の方法。 【請求項9】 Th17細胞の活性化に起因する疾患が、自己免疫疾患、アレルギー疾患又は動脈硬化症である、請求項7又は8記載の方法。 【請求項10】 Pla2g12a、Lplat1、Gpam、Gpat3、及びScd2からなる群から選択されるいずれかの脂質代謝酵素をコードするDNA又はmRNAに生理学的条件下でハイブリダイズし、それによってその転写及び/又は翻訳を阻害する核酸、或いは当該核酸の発現ベクターの有効量を含む、Th17細胞の活性化に起因する疾患の予防又は治療剤。 (【請求項11】以降は省略されています) 発明の詳細な説明【技術分野】 【0001】 本発明は、脂質代謝産物のTh17細胞分化促進剤としての使用に関する。また、本発明は、当該脂質代謝産物やその生合成に関わる脂質代謝酵素を用いたTh17細胞の活性化に起因する疾患の治療薬のスクリーニング方法等に関する。 続きを表示(約 4,300 文字)【背景技術】 【0002】 ナイーブT細胞からTh17細胞への分化は、IL-6やTGFβ等のサイトカインの組み合わせによって制御することができる。Th17細胞は真菌や細胞外細菌に対する宿主の防御に貢献している。しかしながら、Th17細胞は、多発性硬化症、乾癬、炎症性腸疾患、2型糖尿病などの様々な自己免疫疾患の病態に関与していることが、これらの疾患とSTAT3やIL23Rをコードする遺伝子を含むTh17細胞で優先的に発現する遺伝子とを結びつけるゲノムワイド関連研究によって明らかにされている。 【0003】 Th17細胞の分化には、細胞代謝の厳格な制御が必要である。Th17細胞の分化は、de novo脂肪酸生合成の関与に大きく依存している(非特許文献1、2)。Acaca遺伝子によりコードされるアセチル-CoAカルボキシラーゼ1 (ACC1)は、ATP依存的なアセチル-CoAのマロニル-CoAへのカルボキシル化を触媒し、細胞の脂肪酸代謝の第一段階を制御する(非特許文献3)。薬理学的阻害剤である5-テトラデシルオキシ-2-フロイン酸(TOFA)を用いるか、CD4 + T細胞特異的なACC1の欠損によって脂肪酸生合成が阻害されると、Th17細胞の分化が阻害される。さらに、ACC1を欠損させると、IL-17A産生メモリー表現型CD4 + T細胞の割合と数が減少する(非特許文献1)。 【0004】 興味深いことに、脂肪酸フラックスもTh17可塑性の重要な制御因子として浮上してきた(非特許文献1、2、4)。ACC1の薬理学的阻害や遺伝的欠失は、Th17へ偏らせる条件下で培養したCD4 + T細胞において、制御性T細胞に特徴的な転写因子Foxp3の発現を増加させた(非特許文献1、2)。注目すべきことに、このACC1欠損細胞における制御性T細胞分化へのシフトは、外因性脂肪酸(16:0又は18:1)の添加によって克服することができた。このことは、脂肪酸又はその代謝産物が、Th17細胞と制御性T細胞のバランスを調節する上で重要な役割を果たしている可能性を示唆する。これらの試験管内の結果と一致して、TOFAの投与又はACC1の遺伝的欠失は、マウスを実験的自己免疫性脳脊髄炎(EAE)の病態から保護し、中枢神経系におけるIL-17A産生CD4 + T細胞の割合を減少させることが判明した(非特許文献1、2)。さらに、TOFA投与は、食餌誘導性肥満マウスにおいてもEAE病態を改善した。さらに、ACC1の下流で機能する酵素である脂肪酸合成酵素の活性を阻害することによって脂肪酸の生合成を阻害すると、Th17細胞の炎症潜在性が低下する(非特許文献5)。 【0005】 核内ホルモン受容体(NHR)の一つであるレチノイド関連オーファン受容体γt (RORγt)は、Th17細胞の分化誘導に必須な役割を果たすマスター転写因子である(非特許文献6)。以前、本発明者らや他の研究者は、Th17細胞の分化過程において、脂肪酸代謝がRORγt活性と関連することを見出した(非特許文献1、非特許文献7)。RORgとRORγtの2つのアイソフォームがRorc遺伝子から転写される。NHRは幅広い発生、生殖、免疫応答プログラムを指令する転写因子である。NHRは、N末端のDNA結合ドメインとC末端のリガンド結合ドメインから構成される共通のモジュール構造を共有している。トランス活性化のためには、ほとんどのNHRはリガンド結合ドメインとリガンドとの相互作用を必要とする。ホルモン、ビタミン、ステロイド、レチノイド、脂肪酸など、いくつかのクラスの親油性低分子がNHRリガンドとして同定されている(非特許文献8)。オーファンNHRの天然リガンドを同定することは、これらの受容体が食事因子や内因性代謝産物によってどのように制御されるかを理解する上で重要なステップである。RORγtは3つのドメインからなる核内受容体であり(非特許文献9、10)、これには、DNA結合を担う2つのジンクフィンガーモチーフを持つ保存されたDNA結合ドメインと、ステロイド受容体転写共活性化因子のファミリーを動員して遺伝子発現を刺激するカルボキシ末端AF2モチーフを持つ保存されたリガンド結合ドメインが含まれる(非特許文献9、11)。 【0006】 シングルセルRNA配列決定(scRNA-seq)アプローチにより、Th17細胞の分化と機能を制御するRORγtのような因子についての理解がより深まってきた。実際、EAEマウスのT細胞のscRNA-seqにより、低レベルのCD5分子ライク(CD5L)発現がTh17の病原性と相関していることが示された(非特許文献7、12)。CD5Lはde novo脂肪酸生合成経路を介してフラックスを制御している。CD5L欠損Th17細胞のリピドーム解析から、リン脂質と中性脂質の両方の脂肪酸組成が全体的に異なり、細胞内の飽和脂肪酸及び一価不飽和脂肪酸のレベルが増加し、アラキドン酸等の多価不飽和脂肪酸が対応して減少することが明らかになった(非特許文献7)。さらに、Th17細胞に外因性の多価不飽和脂肪酸を加えると、Il17及びIl23r遺伝子座におけるRORγtの結合が減少するのに対し、外因性の飽和脂肪酸又は一価不飽和脂肪酸を与えると、標的遺伝子座へのRORγt結合が増加した(非特許文献7)。したがって、脂肪酸組成の細胞内バランスは、少なくとも部分的には、Th17シグネチャーサイトカインに関連する遺伝子座におけるRORγtの動員及び/又は活性を介して、Th17の機能に影響を与えているようである。 【0007】 転写因子の脂質修飾を介したTh17細胞の分化における脂肪酸の重要な役割が報告されている (非特許文献13)。その一例がSTAT3で、パルミトイルトランスフェラーゼDHHC7を介したパルミトイル化がSTAT3の膜への動員を促進し、その活性化を促進する(非特許文献13)。一旦活性化されると、リン酸化STAT3は核内に入り、Th17細胞分化に必要な多くの重要遺伝子を転写活性化する(非特許文献13)。 【0008】 脂肪酸代謝に加えて、コレステロール生合成経路を介した代謝フラックスもTh17細胞の分化に関与しており、RORγt活性の制御に影響を及ぼす可能性がある(非特許文献14~16)。コレステロール生合成経路の遠位枝にある特定の酵素を遺伝的に欠失させると、CD4 + T細胞によるIL-17産生が減少する(非特許文献14)。さらに、コレステロール生合成経路の3-ヒドロキシ-3-メチルグルタリルCoAリダクターゼの薬理学的阻害剤であるスタチンでTh17細胞を処理すると、Th17細胞と制御性T細胞のバランスに影響を与える(非特許文献17)。ラノステロールを含むオキシステロールやコレステロール生合成中間体はRORγt活性を増加させるが(非特許文献14~16)、Th17細胞の分化過程において、どのステロール代謝産物がRORγtの生理学的リガンドとして作用するかは、依然として不明である。興味深いことに、コレステロール生合成の薬理学的阻害も遺伝的欠失も、IL-17産生を完全に抑止することはできなかった(非特許文献14、17)。脂質のホメオスタシスとRORγtの機能を結びつける分子機序は完全には明らかにされていないが、これらの従前の知見は、脂肪酸とコレステロールの代謝産物が、RORγtの生理学的モジュレーターとして共に機能する可能性を示唆している。 【先行技術文献】 【非特許文献】 【0009】 Y. Endo et al., Cell Rep 12, 1042-1055 (2015) L. Berod et al., Nat Med 20, 1327-1333 (2014) S. J. Wakil et al., Annu Rev Biochem 52, 537-579 (1983) D. A. Nicholas et al., Cell Metab 30, 447-461 e445 (2019) K. E. Young et al., J Leukoc Biol 102, 1229-1235 (2017) Ivanov, II et al., Cell 126, 1121-1133 (2006) C. Wang et al., Cell 163, 1413-1427 (2015) P. Huang et al., Annu Rev Physiol 72, 247-272 (2010) A. M. Jetten, Nucl Recept Signal 7, e003 (2009) S. Rutz et al., Cytokine Growth Factor Rev 30, 1-17 (2016) Z. He et al., Nat Immunol 18, 1128-1138 (2017) J. T. Gaublomme et al., Cell 163, 1400-1412 (2015) M. Zhang et al., Nature 586, 434-439 (2020) F. R. Santori et al., Cell Metab 21, 286-298 (2015) X. Hu et al., Nat Chem Biol 11, 741 (2015) P. Soroosh et al., Proc Natl Acad Sci U S A 111, 12163-12168 (2014) S. Kagami et al., Int Immunol 21, 679-689 (2009) 【発明の概要】 【発明が解決しようとする課題】 【0010】 本発明は、Th17細胞分化を制御する脂質代謝産物及びそれに関わる脂質生合成酵素を同定し、それに基づき、過剰なTh17細胞応答に起因する疾患の治療標的を提供することを目的とする。 【課題を解決するための手段】 (【0011】以降は省略されています) この特許をJ-PlatPatで参照する

 特許ウォッチ
特許ウォッチ