TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025063027
公報種別
公開特許公報(A)
公開日
2025-04-15
出願番号
2024214754,2023017919
出願日
2024-12-09,2015-06-18
発明の名称
ドコサペンタエン酸を含む脂質
出願人
コモンウェルス サイエンティフィック アンド インダストリアル リサーチ オーガナイゼーション
,
グレインズ リサーチ アンド デヴェロップメント コーポレイション
,
ヌシード ニュートリショナル オーストラリア ピーティーワイ リミテッド
代理人
個人
,
個人
,
個人
,
個人
主分類
A01H
1/00 20060101AFI20250408BHJP(農業;林業;畜産;狩猟;捕獲;漁業)
要約
【課題】ドコサペンタエン酸(DPA)を含む抽出植物脂質または微生物脂質、及び該抽出脂質の製造方法を提供する。
【解決手段】エステル化された形態の脂肪酸を含み、前記脂肪酸は、オレイン酸、パルミチン酸、リノール酸(LA)を含むω6脂肪酸、α-リノレン酸(ALA)を含むω3脂肪酸及びドコサペンタエン酸(DPA)を含み、場合により、ステアリドン酸(SDA)、エイコサペンタエン酸(EPA)、及びエイコサテトラエン酸(ETA)のうち1つ以上を含んでもよく、前記抽出脂質の総脂肪酸含有量におけるDPAのレベルが、7%から35%の間である、抽出植物脂質または微生物脂質が提供される。
【選択図】なし
特許請求の範囲
【請求項1】
エステル化された形態の脂肪酸を含み、前記脂肪酸は、オレイン酸、パルミチン酸、リ
ノール酸(LA)を含むω6脂肪酸、α-リノレン酸(ALA)を含むω3脂肪酸及びド
コサペンタエン酸(DPA)を含み、場合により、ステアリドン酸(SDA)、エイコサ
ペンタエン酸(EPA)、及びエイコサテトラエン酸(ETA)のうち1つ以上を含んで
もよく、前記抽出脂質の総脂肪酸含有量におけるDPAのレベルが、7%から35%の間
である、抽出植物脂質または微生物脂質。
続きを表示(約 7,100 文字)
【請求項2】
以下の特徴:
i)前記抽出脂質の総脂肪酸含有量におけるパルミチン酸のレベルは、約2%から15
%の間、または約3%から10%の間であること、
ii)前記抽出脂質の総脂肪酸含有量におけるミリスチン酸(C14:0)のレベルは
、約0.1%であること、
iii)該抽出脂質の総脂肪酸含有量におけるオレイン酸のレベルは、約1%から約3
0%の間、約3%から約30%の間、約6%から約30%の間、約1%から約20%の間
、約30%から約60%の間、約45%~約60%、約30%、または約15%から約3
0%の間であること、
iv)該抽出脂質の総脂肪酸含有量におけるリノール酸(LA)のレベルは、約4%か
ら約35%の間、約4%から約20%の間、約4%から約17%の間、または約5%から
約10%の間であること、
v)該抽出脂質の総脂肪酸含有量におけるα-リノレン酸(ALA)のレベルは、約4
%から約40%の間、約7%から約40%の間、約10%から約35%の間、約20%か
ら約35%の間、約4%から約16%の間、または約2%から約16%の間であること、
vi)前記抽出脂質の総脂肪酸含有量におけるγ-リノレン酸(GLA)のレベルは、
4%未満、約3%未満、約2%未満、約1%未満、約0.5%未満、0.05%から7%
の間、0.05%から4%の間、0.05%から約3%の間、または0.05%から約2
%の間であること、
vii)該抽出脂質の総脂肪酸含有量におけるステアリドン酸(SDA)のレベルは、
約10%未満、約8%未満、約7%未満、約6%未満、約4%未満、約3%未満、約0.
05%から約7%の間、約0.05%から約6%の間、約0.05%から約4%の間、約
0.05%から約3%の間、約0.05%から約10%の間、または約0.05%から約
2%の間であること、
viii)該抽出脂質の総脂肪酸含有量におけるエイコサテトラエン酸(ETA)のレ
ベルは、約6%未満、約5%未満、約4%未満、約1%未満、約0.5%未満、0.05
%から約6%の間、0.05%から約5%の間、0.05%から約4%の間、0.05%
から約3%の間、または0.05%から約2%の間であること、
ix)該抽出脂質の総脂肪酸含有量におけるエイコサトリエン酸(ETrA)のレベル
は、4%未満、約2%未満、約1%未満、0.05%から4%の間、0.05%から3%
の間、または0.05%から約2%の間、または0.05%から約1%の間であること、
x)該抽出脂質の総脂肪酸含有量におけるエイコサペンタエン酸(EPA)のレベルは
、4%から15%の間、4%未満、約3%未満、約2%未満、0.05%から10%の間
、0.05%から5%の間、0.05%から約3%の間、または0.05%から約2%の
間であること、
xi)もし、前記抽出脂質の総脂肪酸含有量におけるDHAのレベルが、2%未満、ま
たは0.05%から約2%の間であること、
xii)該脂質は、その脂肪酸含有物中にω6-ドコサペンタエン酸(22:5
Δ4,
7,10,13,16
)を含むこと、
xiii)該脂質は、その脂肪酸含有物中に0.1%未満のω6-ドコサペンタエン酸
(22:5
Δ4,7,10,13,16
)を含むこと、
xiv)該脂質は、その脂肪酸含有物中に0.1%未満の1つ以上または全てのSDA
、EPA及びETAを含むこと、
xv)該抽出脂質の総脂肪酸含有量における総飽和脂肪酸レベルは、約4%から約25
%の間、約4%から約20%の間、約6%から約20%の間、または約6%から約12%
の間であること、
xvi)該抽出脂質の総脂肪酸含有量における総一価不飽和脂肪酸レベルは、約4%か
ら約40%の間、約4%から約35%の間、約8%から約25%の間、8%から約22%
【請求項3】
前記脂質がオイル、好ましくは、油料種子から採れるオイルであり、より好ましくは、
前記脂質が、セイヨウアブラナ(Brassica napus)オイルまたはカラシナ
(Brassica juncea)オイルなどのアブラナ属(Brassica sp
.)オイル、ワタ(Gossypium hirsutum)オイル、アマ(Linum
usitatissimum)オイル、ヘリアンタス属(Helianthus sp
.)オイル、ベニバナ(Carthamus tinctorius)オイル、ダイズ(
Glycine max)オイル、トウモロコシ(Zea mays)オイル、ギニアア
ブラヤシ(Elaesis guineenis)オイル、ベンサミアナタバコ(Nic
otiana benthamiana)オイル、アオバナルーピン(Lupinus
angustifolius)オイル、カメリナサティバ(Camelina sati
va)オイル、クランベアビシニカ(Crambe abyssinica)オイル、ジ
ャイアントミスカンサス(Miscanthus x giganteus)オイル、ま
たはススキ(Miscanthus sinensis)オイルを含むまたはである、請
求項1または2のいずれか1項に記載の脂質。
【請求項4】
i)脂質を含む植物部分または微生物細胞を得る工程であって、前記脂質がエステル化
された形態の脂肪酸を含み、前記脂肪酸がオレイン酸、パルミチン酸、リノール酸(LA
)を含むω6脂肪酸、α-リノレン酸(ALA)を含むω3脂肪酸、ステアリドン酸(S
DA)、ドコサペンタエン酸(DPA)を含み、場合により、1つ以上のエイコサペンタ
エン酸(EPA)及びエイコサテトラエン酸(ETA)を含んでもよく、前記植物部分中
の抽出脂質の総脂肪酸含有量におけるDPAのレベルが7%から35%の間である前記工
程、及び
ii)前記植物部分または微生物細胞から脂質を抽出する工程
を含み、前記抽出脂質の総脂肪酸含有量におけるDPAのレベルが7%から35%の間で
ある抽出植物脂質または微生物脂質の産生方法。
【請求項5】
前記抽出脂質が請求項2または請求項3で定義された特徴の1つ以上を有する、請求項
4記載の方法
【請求項6】
前記植物部分が種子、好ましくは、セイヨウアブラナ(Brassica napus
)またはカラシナ(Brassica juncea)などのアブラナ属(Brassi
ca sp.)、ワタ(Gossypium hirsutum)、アマ(Linum
usitatissimum)、ヘリアンタス属(Helianthus sp.)、ベ
ニバナ(Carthamus tinctorius)、ダイズ(Glycine ma
x)、トウモロコシ(Zea mays)、ギニアアブラヤシ(Elaesis gui
neenis)、ベンサミアナタバコ(Nicotiana benthamiana)
、アオバナルーピン(Lupinus angustifolius)、カメリナサティ
バ(Camelina sativa)、またはクランベアビシニカ(Crambe a
byssinica)、好ましくは、セイヨウアブラナ(Brassica napus
)、カラシナ(B. juncea)またはカメリナサティバ(C. sativa)の
種子などの油料種子である、請求項4または5に記載の方法。
【請求項7】
前記植物部分または微生物細胞が、酵素の以下のセット:
i)ω3-デサチュラーゼ、Δ6-デサチュラーゼ、Δ5-デサチュラーゼ、Δ6-エ
ロンガーゼ及びΔ5-エロンガーゼ、
ii)Δ15-デサチュラーゼ、Δ6-デサチュラーゼ、Δ5-デサチュラーゼ、Δ6
-エロンガーゼ及びΔ5-エロンガーゼ、
iii)Δ12-デサチュラーゼ、Δ6-デサチュラーゼ、Δ5-デサチュラーゼ、Δ
6-エロンガーゼ及びΔ5-エロンガーゼ、
iv)Δ12-デサチュラーゼ、ω3-デサチュラーゼ及び/またはΔ15-デサチュ
ラーゼ、Δ6-デサチュラーゼ、Δ5-デサチュラーゼ、Δ6-エロンガーゼ及びΔ5-
エロンガーゼ、
v)ω3-デサチュラーゼ、Δ8-デサチュラーゼ、Δ5-デサチュラーゼ、Δ9-エ
ロンガーゼ及びΔ5-エロンガーゼ、
vi)Δ15-デサチュラーゼ、Δ8-デサチュラーゼ、Δ5-デサチュラーゼ、Δ9
-エロンガーゼ及びΔ5-エロンガーゼ、
vii)Δ12-デサチュラーゼ、Δ8-デサチュラーゼ、Δ5-デサチュラーゼ、Δ
9-エロンガーゼ及びΔ5-エロンガーゼ、
viii)Δ12-デサチュラーゼ、ω3-デサチュラーゼ及び/またはΔ15-デサ
チュラーゼ、Δ8-デサチュラーゼ、Δ5-デサチュラーゼ、Δ9-エロンガーゼ及びΔ
5-エロンガーゼ、
ix)ω3-デサチュラーゼまたはΔ15-デサチュラーゼ、Δ6-デサチュラーゼ、
Δ5-デサチュラーゼ、Δ6-エロンガーゼ及びΔ5-エロンガーゼ、または
x)ω3-デサチュラーゼまたはΔ15-デサチュラーゼ、Δ8-デサチュラーゼ、Δ
5-デサチュラーゼ、Δ9-エロンガーゼ及びΔ5-エロンガーゼ
の1つをコードする外来性ポリヌクレオチドを含み、各ポリヌクレオチドは、前記植物部
分または微生物細胞の細胞内の前記ポリヌクレオチドの発現を誘導可能である1つ以上の
プロモーターと作動可能に結合している、請求項4~6のいずれか1項に記載の方法。
【請求項8】
前記植物部分または微生物細胞が、以下の特徴:
i)前記Δ12‐デサチュラーゼが、少なくとも約60%、少なくとも約70%、少な
くとも約80%、約60%から約95%の間、約70%から約90%の間、または約75
%から約85%の間の効率で、前記植物部分または微生物細胞の1つ以上の細胞内で、オ
レイン酸をリノール酸に転換すること、
ii)前記ω3‐デサチュラーゼが、少なくとも約65%、少なくとも約75%、少な
くとも約85%、約65%から約95%の間、約75%から約91%の間、または約80
%から約91%の間の効率で、前記植物部分または微生物細胞の1つ以上の細胞内で、ω
6脂肪酸をω3脂肪酸に転換すること、
iii)前記Δ6‐デサチュラーゼが、少なくとも約20%、少なくとも約30%、少
なくとも約40%、少なくとも約50%、少なくとも約60%、少なくとも約70%、約
30%から約70%の間、約35%から約60%の間、または約50%から約70%の間
の効率で、前記植物部分または微生物細胞の1つ以上の細胞内で、ALAをSDAに転換
すること、
iv)前記Δ6‐デサチュラーゼが、約5%未満、約2.5%未満、約1%未満、約0
.1%から約5%の間、約0.5%から約2.5%の間、または約0.5%から約1%の
間の効率で、前記植物部分または微生物細胞の1つ以上の細胞内で、リノール酸をγ‐リ
ノレン酸に転換すること、
v)前記Δ6‐エロンガーゼが、少なくとも約60%、少なくとも約70%、少なくと
も約75%、約60%から約95%の間、約70%から約80%の間、または約75%か
ら約80%の間の効率で、前記植物部分または微生物細胞の1つ以上の細胞内で、SDA
をETAに転換すること、
vi)前記Δ5‐デサチュラーゼが、少なくとも約60%、少なくとも約70%、少な
くとも約75%、少なくとも約80%、少なくとも約90%、約60%から約95%の間
、約70%から約95%の間、または約75%から約95%の間の効率で、前記植物部分
または微生物細胞の1つ以上の細胞内で、ETAをEPAに転換すること、
vii)前記Δ5‐エロンガーゼが、少なくとも約80%、少なくとも約85%、少な
くとも約90%、約50%から約90%の間、または約85%から約95%の間の効率で
、前記植物部分または微生物細胞の1つ以上の細胞内で、EPAをDPAに転換すること
、
viii)前記植物部分または微生物細胞の1つ以上の細胞内で、オレイン酸をDPA
に転換する効率が、少なくとも約10%、少なくとも約15%、少なくとも約20%、少
なくとも約25%、約20%、約25%、約30%、約10%から約50%の間、約10
%から約30%の間、または約10%から約25%の間、または約20%から約30%の
間であること、
ix)前記植物部分または微生物細胞の1つ以上の細胞内で、LAをDPAに転換する
効率が、少なくとも約15%、少なくとも約20%、少なくとも約22%、少なくとも約
25%、少なくとも約30%、約25%、約30%、約35%、約15%から約50%の
間、約20%から約40%の間、または約20%から約30%の間であること、
x)前記植物部分または微生物細胞の1つ以上の細胞内で、ALAをDPAに転換する
効率が、少なくとも約17%、少なくとも約22%、少なくとも約24%、少なくとも約
30%、約30%、約35%、約40%、約17%から約55%の間、約22%から約3
5%の間、または約24%から約35%の間であること、
xi)前記植物部分または微生物細胞の1つ以上の細胞は、前記外来性ポリヌクレオチ
ドを含まない対応細胞より、少なくとも約25%、少なくとも約30%、約25%から約
40%の間、または約27.5%から約37.5%の間で多いω3脂肪酸を含むこと、
xii)前記Δ6-デサチュラーゼは、リノール酸(LA)と比較してα-リノレン酸
(ALA)を優先的に不飽和化すること、
xiii)前記Δ6-エロンガーゼは、Δ9-エロンガーゼ活性も有すること、
xiv)前記Δ12-デサチュラーゼは、Δ15-デサチュラーゼ活性も有すること、
【請求項9】
前記植物部分または微生物細胞が、ジアシルグリセロールアシルトランスフェラーゼ(
DGAT)、モノアシルグリセロールアシルトランスフェラーゼ(MGAT)、グリセロ
ール-3-リン酸アシルトランスフェラーゼ(GPAT)、1-アシル-グリセロール-
3-リン酸アシルトランスフェラーゼ(LPAAT)、好ましくは、DPA-CoAなど
のC22多価不飽和脂肪アシル-CoA基質を使用できるLPAAT、アシル-CoA:
リゾホスファチジルコリンアシルトランスフェラーゼ(LPCAT)、ホスホリパーゼA
2
(PLA
2
)、ホスホリパーゼC(PLC)、ホスホリパーゼD(PLD)、CDP-
コリンジアシルグリセロールコリンホスホトランスフェラーゼ(CPT)、ホスファチジ
ルコリンジアシルグリセロールアシルトランスフェラーゼ(PDAT)、ホスファチジル
コリン:ジアシルグリセロールコリンホスホトランスフェラーゼ(PDCT)、アシル-
CoAシンターゼ(ACS)、またはその2つ以上の組合せをコードする外来性ポリヌク
レオチドをさらに含む、請求項7または請求項8記載の方法。
【請求項10】
前記外来性ポリヌクレオチドが、前記植物部分または微生物細胞の細胞のゲノムに組み
込まれたDNA分子、好ましくは、T-DNA分子内に共有結合しており、好ましくは、
前記植物部分または微生物細胞の細胞のゲノムに組み込まれたかかるDNA分子数が、1
以下、2もしくは3以下であり、または2もしくは3である、請求項7~9のいずれか1
項に記載の方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、植物細胞または微生物細胞から得られたドコサペンタエン酸を含む脂質、及
び該脂質の産生及び使用方法に関する。
続きを表示(約 5,500 文字)
【背景技術】
【0002】
ω3長鎖多価不飽和脂肪酸(LC-PUFA)は、ヒト及び動物の健康のための重要な
化合物として、今では広く認識されている。これらの脂肪酸を、食事供給源からまたは、
その両方とも食餌の必須脂肪酸として見なされているリノール酸(LA、18:2ω6)
またはα-リノレン酸(ALA、18:3ω3)脂肪酸の転換により得ることができる。
ヒト及び多くの他脊椎動物は、植物源から得られたLAまたはALAをC22に転換でき
るが、この転換反応は、非常に遅い速度で起こる。さらに、現代社会のほとんどは、理想
とされるω6:ω3脂肪酸について4:1以下の比の代わりに、少なくとも90%の多価
不飽和脂肪酸(PUFA)がω6脂肪酸であるというバランスのとれていない食事をとっ
ている(Trautwein、2001)。ヒトのためのエイコサペンタエン酸(EPA
、20:5ω3)、ドコサペンタエン酸(DPA)、及びドコサヘキサエン酸(DHA、
22:6ω3)などのLC-PUFAの即時型食事供給源は、ほとんど、魚または魚油か
らである。従って、健康専門家はヒトの食事に、LC-PUFAの有意なレベルを含む魚
を規則的に含めることを推奨している。益々、魚由来LC-PUFAオイルは、例えば、
食品及び乳児用調製粉乳に配合されている。しかしながら、世界的及び国の水産業の衰退
のせいで、これらの有益な健康増進オイルの代替源が必要とされている。
【0003】
動物と対照的に、顕花植物類は、18個の炭素より長い長鎖を有する多価不飽和脂肪酸
を合成する能力を有しない。特に、他被子植物類と共に作物及び園芸植物は、ALAから
誘導されるEPA、ドコサペンタエン酸(DPA、22:5ω3)及びDHAなどのより
長鎖ω3脂肪酸を合成するために必要な酵素を有しない。従って、植物バイオ技術の重要
ゴールは、実質的量のLC-PUFAを産生し、それ故、これらの化合物の代替供給源を
提供する作物のエンジニアリングである。
【発明の概要】
【0004】
LC-PUFA生合成経路
微細藻類、セン類及び真菌類などの生物のLC-PUFAの生合成は、通常、一連の酸
素依存性不飽和化及び伸長反応として起こる(図1)。これらの生物内でEPAを産生す
る最も共通の経路としては、Δ6-不飽和化反応、Δ6-伸長反応及びΔ5-不飽和化反
応(Δ6-不飽和化反応経路と呼ばれる)が挙げられ、一方で、余り共通でない経路は、
Δ9-伸長反応、Δ8-不飽和化反応及びΔ5-不飽和化反応(Δ9-不飽和化反応経路
と呼ばれる)を使用する。これらの連続した不飽和化及び伸長反応は、図1の上左部に模
式的に示したω6脂肪酸基質LA(ω6)または図1の下右部に示したEPAに完結する
ω3基質ALA(ω3)のいずれかで開始し得る。もし、最初のΔ6-不飽和化反応が、
ω6基質LA上で起こるならば、一連の3種の酵素のLC-PUFA産生物は、ω6脂肪
酸ARAであろう。LC-PUFA合成生物は、アラキドン酸(ARA、20:4ω6)
のEPAへの転換について図1中のΔ17-デサチュラーゼ工程として示したω3-デサ
チュラーゼを用いて、ω6脂肪酸をω3脂肪酸に転換し得る。ω3-デサチュラーゼファ
ミリーのいくつかのメンバーは、LAからARAまでの範囲の様々な基質上で作用できる
。植物ω3-デサチュラーゼは、しばしば、LAからALAまでのΔ15-不飽和化反応
を特異的に触媒するが、一方、真菌類及び酵母のω3-デサチュラーゼは、ARAからE
PAまでのΔ17-不飽和化反応に対して特異的であり得る(Pereiraら,200
4a;Zankら,2005)。広範囲のω6基質をそれらの対応するω3産生物に転換
できる非特異的ω3-デサチュラーゼが存在し得ることを、いくつかの報告が示唆してい
る(Zhangら,2008)。
【0005】
これらの生物内におけるEPAのDHAへの転換は、EPAのΔ5-伸長反応により起
こり、DPAを産生し、次いで、Δ4-不飽和化反応によりDHAを産生する(図1)。
対照的に、哺乳類は、Δ4-デサチュラーゼと独立した3種の別反応によりDPAをDH
Aに転換する、いわゆる、「シュプレッヒャー」経路を使用する(Sprecherら,
1995)。
【0006】
植物、セン類、微細藻類、及び線虫(Caenorhabditis elegans
)などの下等動物で概して見られるフロントエンドデサチュラーゼは、ホスファチジルコ
リン(PC)基質のsn-2位とエステル化された脂肪酸基質を支配的にに受容する。従
って、これらのデサチュラーゼは、アシル-PC、脂質結合、フロントエンドデサチュラ
ーゼとして知られる(Domergueら,2003)。対照的に、高等動物フロントエ
ンドデサチュラーゼは、概して、脂肪酸基質がPCよりCoAと結合しているアシル-C
oA基質を受容する(Domergueら,2005)。いくつかの微細藻類デサチュラ
ーゼ及び1つの植物デサチュラーゼは、CoAとエステル化された脂肪酸基質を使用する
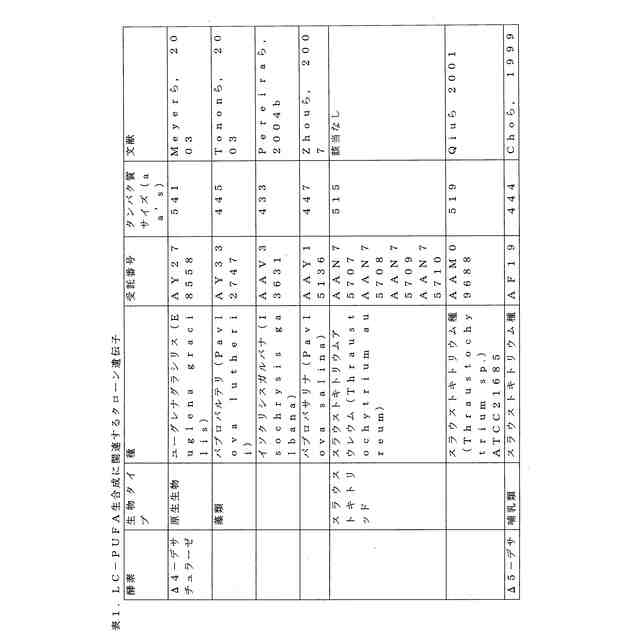
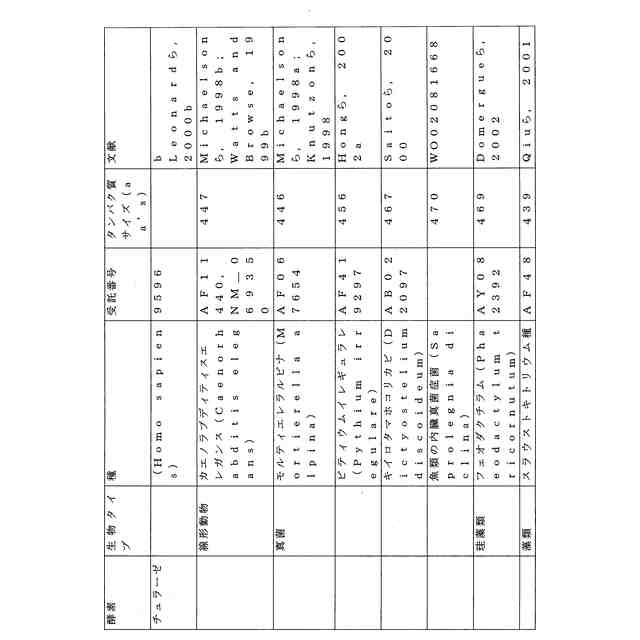
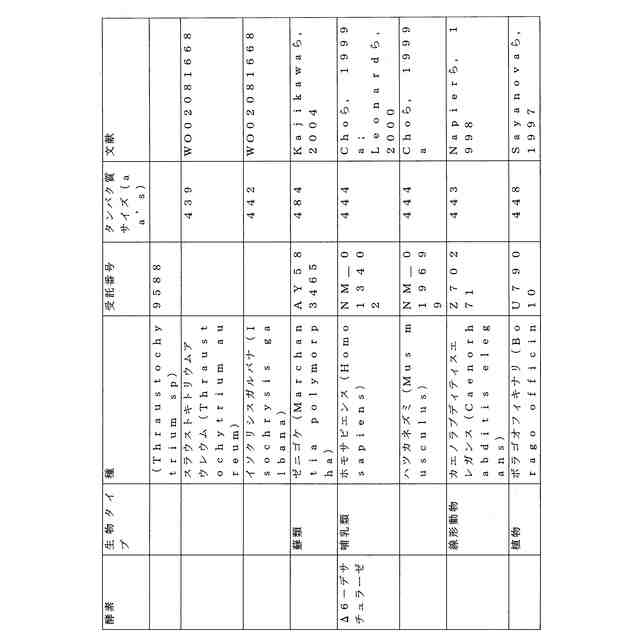
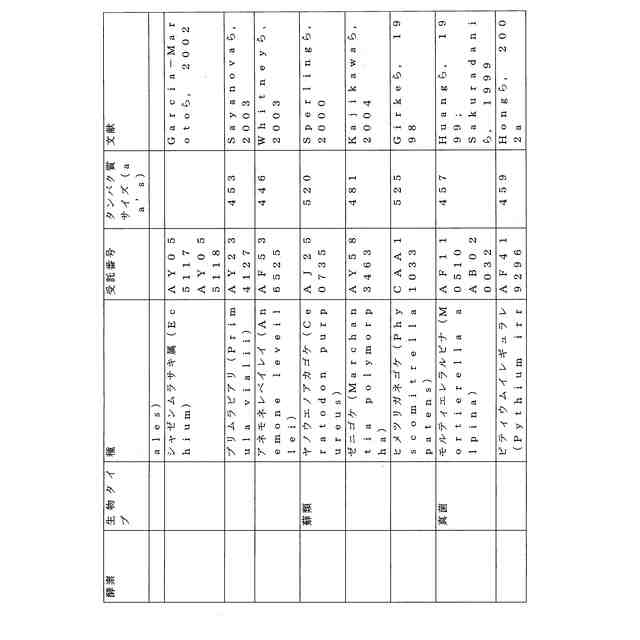
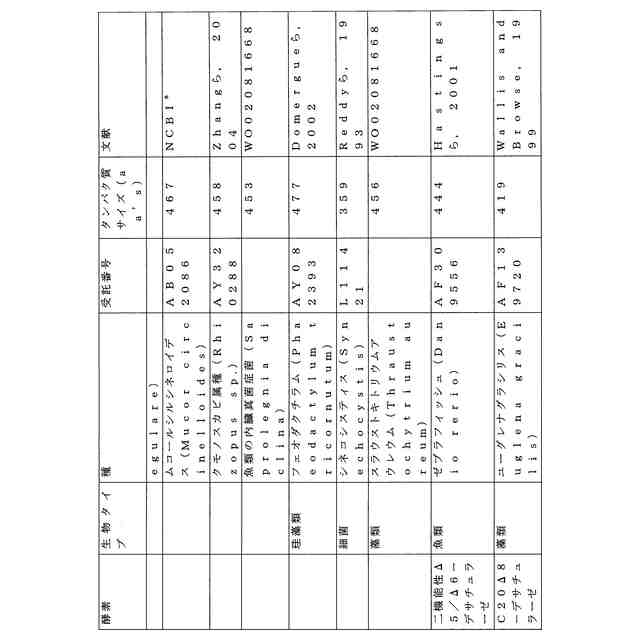
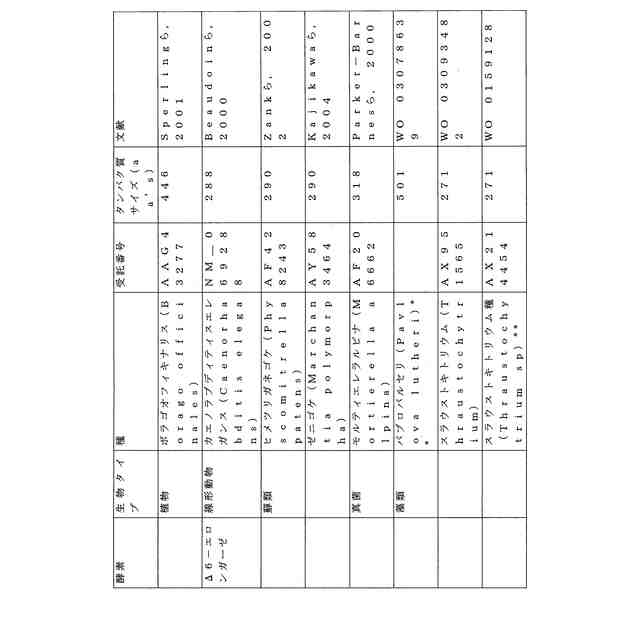
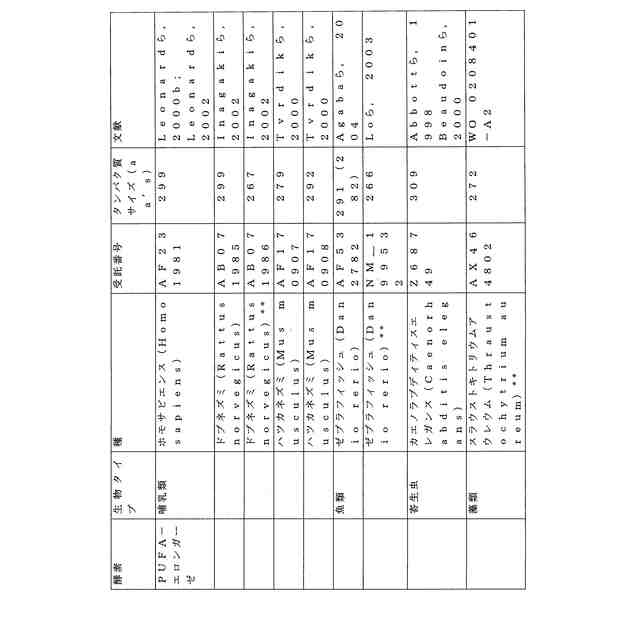
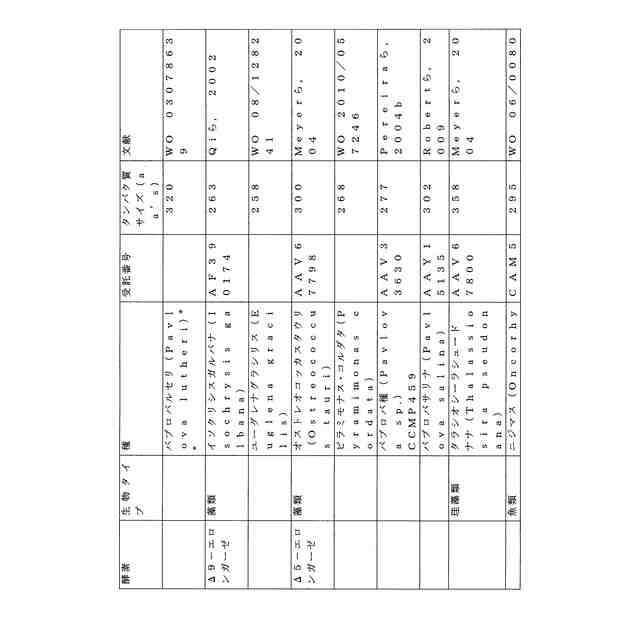
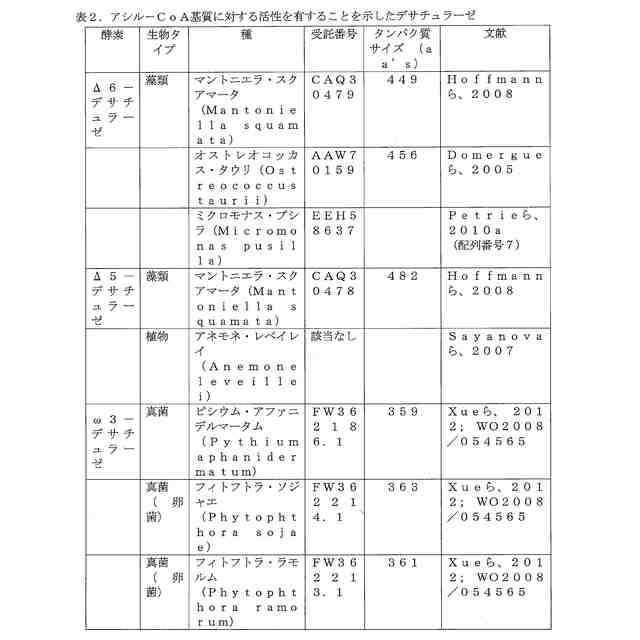
ことが知られている(表2)。
【0007】
各PUFA伸長反応は、他成分タンパク質複合体により触媒される4工程から成る:先
ず、縮合反応の結果、マロニル-CoAから脂肪酸に2C単位が付加され、β-ケトアシ
ル中間体を生成する。それから、これが、NADPHにより還元され、次いで、脱水反応
によりエノイル中間体を得る。この中間体は、最終的に、2回還元されて、伸長脂肪酸を
生成する。他工程はそうでないが、これらの4反応の縮合工程は基質特異的であると、概
して、考えられる。実際に、天然植物伸長機構は、非天然PUFA基質の伸長において天
然植物伸長機構の効率が低くあり得るが、PUFAに特異的縮合酵素(通常、「エロンガ
ーゼ」と呼ぶ)が導入されること提供するPUFAを伸長可能であることを、このことは
意味している。2007年に、酵母伸長サイクルデヒドラターゼの同定及び特徴付けが発
表された(DenicとWeissman、2007)。
【0008】
伸長反応がアシル-CoAプールにおいて基質に対して起こる一方で、植物、セン類及
び微細藻類のPUFA不飽和化は、アシル-PCプールにおいて優先的に脂肪酸基質に対
して自然に起こる。PC担体へのアシル-CoA脂肪酸の転移がリゾホスファチジルコリ
ンアシルトランスフェラーゼ(LPCAT)により起こる一方で、アシル-PC分子から
CoA担体への脂肪酸の転移は、ホスホリパーゼ(PLA)により起こる(Singhら
、2005)。
【0009】
LC-PUFAの工学的産生
ほとんどのLC-PUFA代謝工学は、好気性Δ6-不飽和化/伸長経路を用いて行わ
れている。タバコ中にγ-リノレン酸(GLA、18:3ω6)の生合成は、シアノバク
テリア・シネコシスティス(cyanobacterium Synechocysti
s)のΔ6-デサチュラーゼを用いて、1996年に初めて報告された(ReddyとT
homas、1996)。より近年では、GLAは、ベニバナ(種子オイル中GLA73
%、WO2006/127789)及びダイズ(GLA28%;Satoら、2004)
などの作物で産生された。EPA及びDHAなどのLC-PUFAの産生は、関連する不
飽和化及び伸長工程数が多いせいで、より複雑な工学操作を伴う。EPA陸上植物のEP
A産生は、Qiらにより初めて報告され(2004)、イソクリシス・ガルバナ(Iso
chrysis galbana)のΔ9-エロンガーゼ、ユーグレナ・グラシリス(E
uglena gracilis)のΔ8-デサチュラーゼ及びモルティエレラ・アルピ
ナ(Mortierella alpina)のΔ5-デサチュラーゼをコードする遺伝
子を、EPA3%までの収率で、アラビドプシス属(Arabidopsis)に導入し
た。この研究後、Abbadiら(2004)が続き、ニセツリガネゴケ(Physco
mitrella patens)のΔ6-デサチュラーゼ及びΔ6-エロンガーゼ及び
フェオダクチラム・トリコルヌタム(Phaeodactylum tricornut
um)のΔ5-デサチュラーゼをコードする遺伝子を用いて、亜麻仁でEPA0.8%ま
での産生を報告した。
【0010】
DHA産生の最初の報告は、ダイズ胚芽内におけるDHA3%の産生を記載しているW
O04/017467であったが、種子ではなく、サプロレグニア・ディクリナ(Sap
rolegnia diclina)Δ6-デサチュラーゼ、モルティエレラ・アルピナ
(Mortierella alpina)Δ6-デサチュラーゼ、モルティエレラ・ア
ルピナ(Mortierella alpina)Δ5-デサチュラーゼ、サプロレグニ
ア・ディクリナ(Saprolegnia diclina)Δ4-デサチュラーゼ、サ
プロレグニア・ディクリナ(Saprolegnia diclina)Δ17-デサチ
ュラーゼ、モルティエレラ・アルピナ(Mortierella alpina)Δ6-
エロンガーゼ及びパブロバ・ルセリ(Pavlova lutheri)Δ5-エロンガ
ーゼをコードする遺伝子導入によるものであった。DHAも産生する胚芽内の最大EPA
レベルは19.6%であり、EPAからDHAへの転換効率は小さいことを示した(WO
2004/071467)。この発見は、ゼブラフィッシュ(Danio rerio)
Δ5/6-デサチュラーゼ、線虫(Caenorhabditis elegans)Δ
6-エロンガーゼ、並びにパブロバ・サリナ(Pavlova salina)Δ5-エ
ロンガーゼ及びΔ4-デサチュラーゼを用いて、アラビドプシス属(Arabidops
is)内におけるEPA3%及びDHA0.5%の産生についてRobertら(200
5)により発表されたものと類似していた。また、2005年に、Wuらは、ピシリウム
・イレグラーレ(Pythium irregulare)Δ6-エロンガーゼ、スラウ
ストキトリッド(Thraustochytrid)Δ5-デサチュラーゼ、ニセツリガ
ネゴケ(Physcomitrella patens)Δ6-エロンガーゼ、キンセン
カ(Calendula officianalis)Δ12-デサチュラーゼ、スラウ
ストキトリッド(Thraustochytrid)Δ5-エロンガーゼ、フィトフトラ
・インフェスタンス(Phytophthora infestans)Δ17-デサチ
ュラーゼ、ニジマス(Oncorhyncus mykiss)LC-PUFAエロンガ
ーゼ、スラウストキトリッド(Thraustochytrid)Δ4-デサチュラーゼ
及びスラウストキトリッド(Thraustochytrid)LPCATを用いて、カ
ラシナ(Brassica juncea)内において、ARA25%、EPA15%、
及びDHA1.5%の産生を発表した(Wuら、2005)。ω3LC-PUFAを合成
する油料種子作物を産生する試みの総括が、Venegas-Canelonら(201
0)及びRuiz-Lopezら(2012)により提供されている。 Ruiz-Lo
pezら(2012)により示されたように、トランスジェニック植物内のDHA産生に
ついて、今日までに得られた結果は魚油で見られるレベルにはほど遠いものであった。よ
り近年、Petrieら(2012)は、シロイヌナズナ(Arabidopsis t
haliana)の種子内におけるDHA約15%の産生を報告し、WO2013/18
5184は、7%から20%の間のDHAを有する特定の種子オイルの産生を報告した。
しかしながら、20%を超えるDHAを有する植物油の産生の報告はない。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
個人
昆虫捕獲器
26日前
個人
トラップタワー
25日前
井関農機株式会社
作業車両
26日前
井関農機株式会社
作業車両
1か月前
井関農機株式会社
収穫作業機
1か月前
個人
プランターシーブ
1か月前
株式会社猫壱
首輪
17日前
有限会社小林工業
エビ養殖設備
18日前
個人
ペット用デンタルおもちゃ
25日前
個人
ショベルバケット型スコップ
1か月前
個人
ペット用排泄物収納袋取付具
1か月前
井関農機株式会社
作業車両
17日前
井関農機株式会社
作業車両
24日前
トヨタ自動車株式会社
飛行体
1か月前
井関農機株式会社
農業支援システム
1か月前
株式会社ダイイチ
海苔網の酸処理船
9日前
平岡織染株式会社
防虫間仕切りシート
16日前
株式会社サタケ
穀物タンクの換気装置
10日前
平岡織染株式会社
防虫間仕切りシート
24日前
松山株式会社
草刈作業機
9日前
池田食研株式会社
腹足類の駆除忌避剤
9日前
ニシム電子工業株式会社
散布装置
9日前
東都興業株式会社
シート開閉装置
27日前
株式会社ササキコーポレーション
農作業機
9日前
株式会社ササキコーポレーション
農作業機
9日前
井関農機株式会社
収穫作業機
1か月前
株式会社ササキコーポレーション
農作業機
9日前
第一ビニール株式会社
植物の茎保持具
24日前
第一ビニール株式会社
栽培容器載置台
24日前
株式会社 ジークラック
ルアー
11日前
ポンポリース株式会社
動物用歩行補助具
25日前
株式会社クボタ
水田作業機
2日前
株式会社クボタ
水田作業機
2日前
株式会社シマノ
ルアー
17日前
株式会社アテックス
草刈機の弾性カバー保持機構
9日前
IWKイノベーション株式会社
止まり木ステップ
17日前
続きを見る
他の特許を見る

 特許ウォッチ
特許ウォッチ