TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2023138998
公報種別
公開特許公報(A)
公開日
2023-10-03
出願番号
2023108341,2021108614
出願日
2023-06-30,2014-12-13
発明の名称
DNA抗体構築物及びその使用方法
出願人
ザ トラスティーズ オブ ザ ユニバーシティ オブ ペンシルバニア
,
イノビオ ファーマシューティカルズ,インコーポレイティド
代理人
個人
,
個人
,
個人
,
個人
,
個人
主分類
C12N
15/13 20060101AFI20230926BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】合成抗体またはその断片をインビボ生成する組み換え核酸配列を含む組成物、及び前記組成物を投与することによる対象内の疾患の予防及び/または治療方法を提供すること。
【解決手段】本明細書では、抗体をコードする組み換え核酸配列を含む組成物が開示される。本明細書では、組成物を対象に投与することによる対象内での合成抗体の生成方法も開示される。本開示は、前記組成物及び生成方法を用いて対象内の疾患の予防方法及び/または治療方法も提供する。
【選択図】なし
特許請求の範囲
【請求項1】
(a)配列番号44に記載される核酸配列と;
(b)配列番号67に記載される核酸配列と;
(c)配列番号69に記載される核酸配列と;
(d)配列番号71に記載される核酸配列と;
(e)配列番号73に記載される核酸配列と;
(f)配列番号75に記載される核酸配列と;
(g)配列番号77に記載される核酸配列と;
(h)配列番号58に記載される核酸配列と;
(i)配列番号60に記載される核酸配列と;
(j)配列番号65に記載される核酸配列と
からなる群から選択される核酸配列の全長にわたって少なくとも約95%の同一性を有する前記核酸配列を含む合成抗体をコードする核酸分子。
続きを表示(約 1,500 文字)
【請求項2】
前記核酸配列が、
(a)配列番号44に記載される前記核酸配列と;
(b)配列番号67に記載される前記核酸配列と;
(c)配列番号69に記載される前記核酸配列と;
(d)配列番号71に記載される前記核酸配列と;
(e)配列番号73に記載される前記核酸配列と;
(f)配列番号75に記載される前記核酸配列と;
(g)配列番号77に記載される前記核酸配列と;
(h)配列番号58に記載される前記核酸配列と;
(i)配列番号60に記載される前記核酸配列と;
(j)配列番号65に記載される前記核酸配列と
からなる群から選択される、請求項1に記載の核酸分子。
【請求項3】
(a)配列番号45に記載されるアミノ酸配列と;
(b)配列番号68に記載されるアミノ酸配列と;
(c)配列番号70に記載されるアミノ酸配列と;
(d)配列番号72に記載されるアミノ酸配列と;
(e)配列番号74に記載されるアミノ酸配列と;
(f)配列番号76に記載されるアミノ酸配列と;
(g)配列番号78に記載されるアミノ酸配列と;
(h)配列番号59に記載されるアミノ酸配列と;
(i)配列番号61に記載されるアミノ酸配列と;
(j)配列番号66に記載されるアミノ酸配列と
からなる群から選択されるアミノ酸配列の全長にわたって少なくとも約95%の同一性を有する前記アミノ酸配列を含む合成抗体をコードする核酸分子。
【請求項4】
前記核酸が、
(a)配列番号45に記載される前記アミノ酸配列と;
(b)配列番号68に記載される前記アミノ酸配列と;
(c)配列番号70に記載される前記アミノ酸配列と;
(d)配列番号72に記載される前記アミノ酸配列と;
(e)配列番号74に記載される前記アミノ酸配列と;
(f)配列番号76に記載される前記アミノ酸配列と;
(g)配列番号78に記載される前記アミノ酸配列と;
(h)配列番号59に記載される前記アミノ酸配列と;
(i)配列番号61に記載される前記アミノ酸配列と;
(j)配列番号66に記載される前記アミノ酸配列と
からなる群から選択される前記アミノ酸配列を有するタンパク質をコードする、請求項3に記載の核酸分子。
【請求項5】
前記核酸配列が、軽鎖ポリペプチド、重鎖ポリペプチド、軽鎖ポリペプチド及び重鎖ポリペプチドの両方、またはその断片をコードする、請求項1~4のいずれか1項に記載の核酸分子。
【請求項6】
前記核酸配列が軽鎖ポリペプチド及び重鎖ポリペプチドをコードする場合、前記核酸配列が、プロテアーゼ切断部位もコードする、請求項5に記載の核酸分子。
【請求項7】
前記プロテアーゼ切断部位が、前記軽鎖ポリペプチドと前記重鎖ポリペプチドとの間に位置し、前記プロテアーゼ切断部位が、フリン切断部位及び2Aペプチド配列を含む、請求項6に記載の核酸分子。
【請求項8】
前記核酸分子が、免疫グロブリン(Ig)シグナルペプチドをさらにコードする、請求項1~4のいずれか1項に記載の核酸分子。
【請求項9】
前記Igシグナルペプチドが、IgEシグナルペプチドを含む、請求項8に記載の核酸分子。
【請求項10】
前記核酸分子が、発現ベクターを含む、請求項1~4のいずれか1項に記載の核酸分子。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、合成抗体またはその断片をインビボ生成する組み換え核酸配列を含む組成物、及び前記組成物を投与することによる対象内の疾患の予防及び/または治療方法に関する。
続きを表示(約 15,000 文字)
【背景技術】
【0002】
(関連出願の相互参照)
本願は、2013年12月13日出願の国際出願番号PCT/US2013/075137に対する優先権を主張するものであり、この出願は、参照することでその全体が本明細書に組み入れられる。
【0003】
免疫グロブリン分子は、システイン残基間のジスルフィド結合(S-Sとして示す)によって共有結合された軽(L)鎖及び重(H)鎖を2つずつ含む。重鎖(VH)及び軽鎖(VL)の可変ドメインは、抗体分子の結合部位に寄与する。重鎖定常領域は、3つの定常ドメイン(CH1、CH2、及びCH3)と、(柔軟な)ヒンジ領域とからなる。軽鎖も、定常ドメイン(CL)を有する。重鎖及び軽鎖の可変領域は、4つのフレームワーク領域(FR;FR1、FR2、FR3、及びFR4)と、3つの相補性決定領域(CDR:CDR1、CDR2、及びCDR3)とを含む。従って、これらは非常に複雑な遺伝系であり、インビボ組み立てが困難となっている。
【0004】
標的モノクローナル抗体(mAb)は、過去25年間で最も重要な医療用治療の進歩の1つである。この種の免疫療法は今や、多数の自己免疫疾患、癌及び感染の治療に対して日常的に用いられている。悪性腫瘍に対して、現在用いられている免疫グロブリン(Ig)療法の多くは、腫瘍に向けた細胞毒性化学療法レジメンを併用する。この併用法により全体的な生存率が著しく改善されている。特定の癌に対して用いるmAb製剤が多数認可されており、例えば、非ホジキンリンパ腫治療のためのCD20を標的とするキメラmAbであるリツキサン(リツキシマブ)、ならびにCTLA-4を阻害し、黒色腫及び他の悪性腫瘍の治療に用いられているヒトmAbであるイピリムマブ(エルボイ)が挙げられる。さらに、ベバシズマブ(アバスチン)は、VEGF及び腫瘍新血管形成を標的とし、大腸癌の治療に用いられているもう一つの代表的なヒト化mAbである。悪性腫瘍の治療に最も注目されているmAbは、恐らくHer2/neuを標的とするヒト化製剤であるトラスツズマブ(ハーセプチン)であり、患者の一部の乳癌に対して大きな有効性を持つことが実証されている。さらに、多数のmAbが、自己免疫及び特定の血液疾患の治療に用いられている。
【0005】
癌治療に加えて、ジフテリア、A型及びB型肝炎、狂犬病、破傷風、水痘、及び呼吸器合胞体ウイルス(RSV)などの多くの感染に対する防御効力をポリクローナルIgの受動伝達により調節する。実際、いくつかのポリクローナルIg製剤は、積極的なワクチン接種を介して防御Igを生成するのに十分な時間がない状況下の疾患流行地を旅行している個人において特定の感染性因子に対する一時的な防御をもたらす。さらに、免疫不全の子供において、RSV感染を標的とするmAbであるパリビズマブ(シナジス)は、RSVに対して防御することが臨床的に実証されている。
【0006】
抗体ベースの治療はリスクがないわけではない。1つのかかるリスクは、抗体依存性感染増強(ADE)であり、非中和性抗ウイルスタンパク質が宿主細胞へのウイルス侵入をしやすくする場合に起こり、細胞の感染性の増加につながる。いくつかの細胞は、ウイルスが侵入するのに用いる表面上に通常の受容体を有していない。抗ウイルスタンパク質(すなわち、抗体)は、細胞膜内で、これらの細胞のいくつかが有する抗体Fc受容体に結合する。ウイルスは、抗体の他方の末端で抗原結合部位に結合する。このウイルスは、このメカニズムを使用して、ヒトマクロファージに感染することができ、通常は軽症のウイルス感染を引き起こし、生命に関わるようになる。ADEの最も広く知られている例は、デングウイルス(DENV)による感染の状況で発生する。それは、ある血清型のDENVに以前感染したことがある人が、何カ月または何年も経ってから異なる血清型に感染する場合に観察される。かかる場合では、疾患の臨床経過はより深刻になり、これらの人は、ADEが起こっていない人と比べてより高いウイルス血症を有する。これは、一次(最初の)感染は、子供においてほどんどが軽症の疾患(DF)を引き起こすが、二次感染(後日での再感染)は、子供及び成人の双方において深刻な疾患(DHF及び/またはDSS)と関連付けられやすくなるという観察を説明している。4つの抗原性の異なる血清型のDENV(DENV-1~DENV-4)が存在する。DENVによる感染は、感染している血清型に対して終生免疫をもたらす中和同型免疫グロブリンG(IgG)抗体の産生を誘発する。DENVによる感染は、他の3つの血清型に対してある程度の交差防御免疫ももたらす。中和異型抗体の誘発に加えて、DENVによる感染は、ウイルスを部分的にのみまたは全く中和しない異型抗体も誘発することができる。かかる交差反応性であるが非中和な抗体の産生が、より深刻な二次感染の理由かもしれない。いったん白血球内部へ入ると、ウイルスは検出されないまま複製し、最終的に、深刻な疾患を引き起こす非常に高いウイルス力価を生成する。
【0007】
mAb療法の臨床効力は素晴らしいものがある。しかしながら、この治療アプローチの使用及び普及を制限する問題が残っている。これらの問題のいくつかには、これらの複雑な生物製剤の製造コストが高いことが挙げられ、より広範な人口、とりわけ、大きな影響をもたらし得る発展途上国における使用を限定してしまう。さらに、有効性を達成し維持するためにmAbを反復投与する必要性がしばしばあることもロジスティクス及び患者のコンプライアンスという点で障害になり得る。血清IgGとの競合により治療抗体の低いインビボ有効性を減少させるかまたは除去する新規の抗体が必要とされている。デング、HIV、RSV、及び他のものなどのウイルスにおける抗体依存性感染増強を除去することができる新規の抗体が必要とされている。二重特異性抗体、二官能性抗体、及び抗体カクテルは、治療的または予防的であることを証明することが可能ないくつかの機能を行うために必要である。また、これらの抗体製剤の長期安定性は短いことが多く最適ではない。したがって、当該技術分野では、安全かつコスト効率のよい方法で対象に送達することができる合成抗体分子が必要とされている。
【発明の概要】
【0008】
本発明は、(a)配列番号44に記載される核酸配列と;(b)配列番号67に記載される核酸配列と;(c)配列番号69に記載される核酸配列と;(d)配列番号71に記載される核酸配列と;(e)配列番号73に記載される核酸配列と;(f)配列番号75に記載される核酸配列と;(g)配列番号77に記載される核酸配列と;(h)配列番号58に記載される核酸配列と;(i)配列番号60に記載される核酸配列と;(j)配列番号65に記載される核酸配列とからなる群から選択される核酸配列の全長にわたって少なくとも約95%の同一性を有する核酸配列を含む合成抗体をコードする核酸分子に関する。本発明はさらに、上記の核酸分子をそれを必要とする対象に投与することを含む、対象における疾患の予防方法に関する。本発明はさらに、上記の核酸分子をそれを必要とする対象に投与することを含む、対象における疾患の治療方法に関する。
【0009】
本発明は、(a)配列番号45に記載されるアミノ酸配列と;(b)配列番号68に記載されるアミノ酸配列と;(c)配列番号70に記載されるアミノ酸配列と;(d)配列番号72に記載されるアミノ酸配列と;(e)配列番号74に記載されるアミノ酸配列と;(f)配列番号76に記載されるアミノ酸配列と;(g)配列番号78に記載されるアミノ酸配列と;(h)配列番号59に記載されるアミノ酸配列と;(i)配列番号61に記載されるアミノ酸配列と;(j)配列番号66に記載されるアミノ酸配列とからなる群から選択されるアミノ酸配列の全長にわたって少なくとも約95%の同一性を有するタンパク質をコードする核酸配列を含む合成抗体をコードする核酸分子にも関する。本発明は、上記の核酸分子を含む組成物にも関する。本発明はさらに、上記の核酸分子をそれを必要とする対象に投与することを含む、対象における疾患の予防方法に関する。本発明はさらに、上記の核酸分子をそれを必要とする対象に投与することを含む、対象における疾患の治療方法に関する。
【図面の簡単な説明】
【0010】
実施例1に記載のIgG重鎖をコードする核酸配列を示す。
実施例1に記載のIgG軽鎖をコードする核酸配列を示す。
時間(時)対OD450nm(組織培養上清を1:100に希釈)をプロットしたグラフを示す。
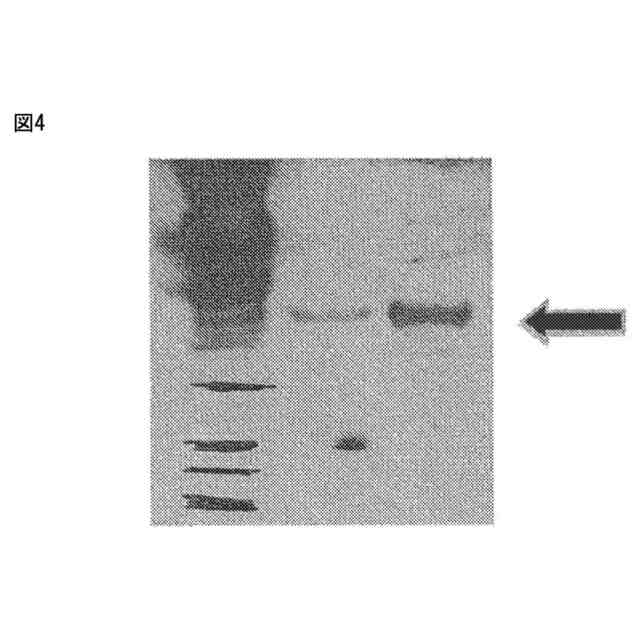
ウエスタンブロットの画像を示す。
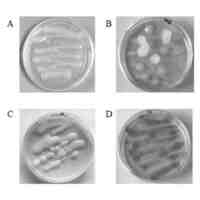
pHIV-1Env-Fabの生成及び発現確認を示す。(A及びB)pHIV-1 Env Fab抗gp120Fab発現構築物の環状プラスミドマップは、VRC01重(H)及び軽(L)可変鎖Ig遺伝子を用いて設計した。発現レベルを増すためにFabプラスミドを構築する際にいくつかの修飾を含んだ。図5に示すように、Fab VL断片遺伝子及びVH断片遺伝子は、pVax1ベクターのBamH1及びXho1制限部位間で別々にクローニングした。(C)pHIV-1 Env Fabのインビトロ発現。グラフは、293T細胞トランスフェクション後のpHIV-1 Env Fab発現の時間的動態を示した。発現を示す値は、3列ウェルの平均値OD450nm±標準偏差である。対照として、293T細胞もpVax1バックボーンでトランスフェクトした。
pHIV-1 Env Fabによる抗HIV Env特異的Fabの時間的生成の測定を示す。(A)抗HIV1 Fabの経時的生成。pHIV-1 Env Fabの投与後、ELISA法によって1:100に最終希釈した血清中で特異的Fabの産生を10日間にわたって測定し、OD450nmとして示した。pVax1投与されたマウスの血清を陰性対照として用いた。(B)組み換えgp120(rgp120)で免疫化した後の抗gp120抗体反応の比較測定。実施例2に記載のように、rgp120を単回注射してマウスを免疫化した後、10日目までの抗gp120抗体の産生を測定し、OD450nm値として示した。この研究において、PBSを陰性対照注射として用いた。(C)免疫ブロット分析によって結合しているHIV1Env-Fabの確認。実施例に記載のように、5μgまたは10μgいずれかのgp120をSDS-PAGE及びニトロセルロースブロッティングに供したした後、pHIV-1 Env Fab投与されたマウスの血清とともにブロットを培養した。実験用血清が結合rgp120を認識したことが免疫ブロットにより示され、生成されたFabの特異性が確認された。(D)ヒトIgG1Fabの時間的定量化。pHIV-1Env-Fabを投与後、マウス血清中のIgG1を測定した。標準ELISAキットにより、示された時点でIgG1を測定し、Fab(μg/mL)±標準偏差で表わした。pVax1投与されたマウスの血清を陰性対照として用いた。x軸上に示された時点で血清サンプルを分析した。図6(A)、(B)、及び(D)に示すグラフの矢印は、DNAプラスミドを投与した時点を示す。
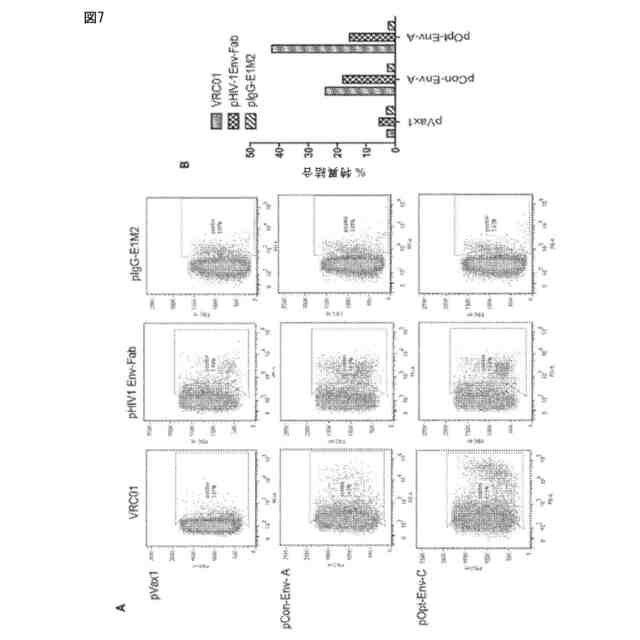
HIV1 Env FabのクレードA HIV Env糖タンパク質へのFACS結合分析を示す。(A)抗HIV1Env-FabのHIV-1クレードA Env糖タンパク質への結合を示すFACSスキャニング。コンセンサス(pCon-Env-A)または「最適化」(pOpt-Env-A)HIV-1クレードAエンベロープのいずれかを発現するDNAを293T細胞にトランスフェクトした。トランスフェクション2日後、精製された天然VRC01 Ig、pHIV-1 Env Fab(プラスミド単回投与後48時間で回収)から生成された血清、またはpIgG-E1M2投与から生成された対照Igのいずれかで細胞を染色した。血清及びVRC01抗体をPBS50μl中でそれぞれ1:4または1:100に希釈し、室温で30分間培養した。その後、適切な二次フィコエリトリン(PE)に複合体化させたIgで細胞を染色し、FACS分析用に単一及び生細胞としてゲートした。陽性細胞の結合率をスキャニングごとに示した。(B)FACS結合データのグラフ図。Ig/血清試験群ごとに染色した細胞数(すなわち、発現レベルを示す)をバックグラウンド染色値で割り、試験した異なるHIVクレードA Env製剤の関数としてy軸上に特異結合率(%)として示した。
pHIV-1Env-Fab投与されたマウスの血清によるHIV-1の経時中和を示す。中和活性の分析に用いた血清は、グラフに示された時点で回収した。中和分析をHIV-1偽型ウイルス:Bal26(パネルA;クレードB、Tier1)、Q23Env17(パネルB;クレードA、Tier1)、SF162S(パネルC;クレードB、Tier1)、及びZM53M(パネルD;クレードC、Tier2)のパネルを用いてTZM-BL細胞で行った。実施例2に記載した通り、0.01のMOIで細胞を感染させ、pHIV-1 Env Fab投与から生成されたFabを含む血清(最終希釈1:50)の存在下で培養した。中和値を%で示す。この計算は実施例2に記載した。なお、グラフごとに示した横線は、実験用血清が50%のウイルス中和を介在したおおよその時点を示す。
実施例2~7に記載のHIV-1 Env Fabの重鎖(VH-CH1)をコードする核酸配列を示す。
実施例2~7に記載のHIV-1 Env Fabの軽鎖(VL-CL)をコードする核酸配列を示す。
HIV Envをコードするプラスミドでトランスフェクトした細胞の免疫蛍光を示す。pVAX1(左パネル)またはpHIV-Env-Fab(右パネル)からの製剤で細胞を染色した。
抗原の種類対血清濃度(ng/mL)をプロットしたグラフを示す。
合成ヒトIgG1抗体をコードする構築物を模式的に示す。
図13の構築物によってコードされる組み立て抗体(発現時)を模式的に示す。
VRC01 IgGのアミノ酸配列を示す。
(A)HIV-1 Env-PG9 Igをコードする構築物の模式図を示す。
(B)(A)の構築物を含むベクターの模式図を示す。
(C)染色したゲルの画像を示す。
(A)HIV-1 Env-4E10 Igをコードする構築物の模式図を示す。
(B)(A)の構築物を含むベクターの模式図を示す。
(C)染色したゲルの画像を示す。
フリンによる切断前のHIV-1 Env-PG9 Igのアミノ酸配列を示す。
フリンによる切断前のHIV-1 Env-4E10 Igのアミノ酸配列を示す。
(A)CHIKV-Env-Fabの重(VH-CH1)鎖をコードする構築物の模式図を示す。
(B)CHIKV-Env-Fabの重(VL-CL)鎖をコードする構築物を模式的に示す。
CHIKV-Env-Fabの重(VH-CH1)または軽(VL-CL)鎖をコードする構築物を含む発現ベクターを模式的に示す。
時単位の時間(hr)対OD450nmをプロットしたグラフを示す。
免疫ブロットの画像を示す。
DNA投与のタイミング、事前採血及び採血を模式的に示す。
日単位の時間対OD450nmをプロットしたグラフを示す。
暴露後の日数対生存率(%)をプロットしたグラフを示す。
マウス群対TNF-αのpg/mLをプロットしたグラフを示す。
マウス群対IL-6のpg/mLをプロットしたグラフを示す。
VH-CH1をコードするプロモーター制御下の構築物を模式的に示す。
VL-CLをコードするプロモーター制御下の構築物を模式的に示す。
発現ベクター内でクローニングした抗Her-2 FabのVH-CH1またはVL-CLをコードする構築物を模式的に示す。
抗Her-2 FabのVH-CH1をコードする核酸配列を示す。
図32の核酸配列によってコードされるアミノ酸配列(すなわち、抗Her-2 FabのVH-CH1のアミノ酸配列)を示す。
抗Her-2 FabのVL-CLをコードする核酸配列を示す。
図34の核酸配列によってコードされるアミノ酸配列(すなわち、抗Her-2 FabのVL-CLのアミノ酸配列)を示す。
トランスフェクト細胞の種類対IgG濃度(μg/mL)をプロットしたグラフを示す。
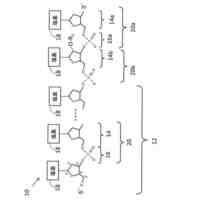
免疫グロブリンG(IgG)重鎖の可変重領域(VH)、可変重定常領域1(CH1)、ヒンジ領域、可変重定常領域2(CH2)、及び可変重定常3(CH3)をコードし、かつ、IgG軽鎖の可変軽領域(VL)及び可変軽定常領域(CL)をコードする構築物を模式的に示す。IgGの重鎖及び軽鎖は、プロテアーゼ切断部位によって分離され、各々は、(リーダー配列によってコードされる)シグナルペプチドによって先行される。
抗デングウイルス(DENV)ヒトIgGをコードする核酸配列を示す。
図39の核酸配列によってコードされるアミノ酸配列(すなわち、抗DENVヒトIgGのアミノ酸配列)を示す。このアミノ酸配列では、重鎖及び軽鎖を2つの別々のポリペプチドに分離させるプロテアーゼ切断がまだ起こっていない。
マウス群対OD450nmをプロットしたグラフを示す。
注射後の日数対ヒトIgG濃度(ng/mL)をプロットしたグラフを示す。
図1の核酸配列(すなわち、配列番号6)によってコードされるアミノ酸配列を示す。このアミノ酸配列は、以下の実施例1に記載のIgG重鎖のアミノ酸配列である。
図2の核酸配列に(すなわち、配列番号7)よってコードされるアミノ酸配列を示す。このアミノ酸配列は、以下の実施例1に記載のIgG軽鎖のアミノ酸配列である。
図9の核酸配列(すなわち、配列番号3)によってコードされるアミノ酸配列を示す。このアミノ酸配列は、実施例2~7に記載のHIV-1 Env-Fabの重鎖(VH-CH1)のアミノ酸配列である。
図10の核酸配列(すなわち、配列番号4)によってコードされるアミノ酸配列を示す。このアミノ酸配列は、実施例2~7に記載のHIV-1 Env-Fabの軽鎖(VL-CL)のアミノ酸配列である。
以下の実施例11に記載のHIV-1 PG9一本鎖Fab(scFab)をコードする核酸配列を示す。
図46の核酸配列(すなわち、配列番号50)によってコードされるアミノ酸配列を示す。このアミノ酸配列は、以下の実施例11に記載のHIV-1 PG9 scFabのアミノ酸配列である。
以下の実施例13に記載のHIV-1 4E10一本鎖Fab(scFab)をコードする核酸配列を示す。
図48の核酸配列(すなわち、配列番号52)によってコードされるアミノ酸配列を示す。このアミノ酸配列は、以下の実施例13に記載のHIV-1 4E10 scFabのアミノ酸配列である。
免疫グロブリンG(IgG)重鎖の可変重領域(VH)、可変重定常領域1(CH1)、ヒンジ領域、可変重定常領域2(CH2)、及び可変重定常3(CH3)をコードする構築物を模式的に示す。IgG重鎖をコードする核酸配列は、リーダー配列によって先行される。
IgG軽鎖の可変軽領域(VL)及び可変軽定常領域(CL)をコードする構築物を模式的に示す。IgG軽鎖をコードする核酸配列は、リーダー配列によって先行される。
以下の実施例9に記載のHIV-1 VRC01 IgG1重鎖をコードする核酸配列を示す。
図52の核酸配列(すなわち、配列番号54)によってコードされるアミノ酸配列を示す。このアミノ酸配列は、以下の実施例9に記載のHIV-1 VRC01 IgG1重鎖のアミノ酸配列である。
以下の実施例9に記載のHIV-1 VRC01 IgG軽鎖をコードする核酸配列を示す。
図54の核酸配列(すなわち、配列番号56)によってコードされるアミノ酸配列を示す。このアミノ酸配列は、以下の実施例9に記載のHIV-1 VRC01 IgG軽鎖のアミノ酸配列である。
以下の実施例14に記載のCHIKV-Env-Fabの重鎖(VH-CH1)をコードする核酸配列を示す。
図56の核酸配列(すなわち、配列番号58)によってコードされるアミノ酸配列を示す。このアミノ酸配列は、以下の実施例14に記載のCHIKV-Env-Fabの重鎖(VH-CH1)のアミノ酸配列である。
以下の実施例14に記載のCHIKV-Env-Fabの軽鎖(VL-CL)をコードする核酸配列を示す。
図58の核酸配列(すなわち、配列番号60)によってコードされるアミノ酸配列を示す。このアミノ酸配列は、以下の実施例14に記載のCHIKV-Env-Fabの軽鎖(VL-CL)のアミノ酸配列である。
以下の実施例12に記載のHIV-1 Env-4E10 Igをコードする核酸配列を示す。
以下の実施例10に記載のHIV-1 Env-PG9 Igをコードする核酸配列を示す。
VRC01 IgG(配列番号64)をコードする核酸配列を示す。
抗体発現プラスミドの概略設計、及びCHIKV-Fab発現プラスミドの単回EP媒介注射後の抗体の発現及び結合速度の確認を示す。(A)選択された抗CHIKVヒトモノクローナルの可変軽鎖及び重鎖(VL及びVH)IgG断片遺伝子を、CHIKV-Fab及びCHIKV-IgGについて別々に、最適化DNAプラスミドベクターにクローニングした。(B)抗CHIKV VL及びVH-Fab遺伝子またはCHIKV-IgGをコードするDNAプラスミドを、ELISA法によってそれぞれのインビトロ発現を決定するために、293T細胞に一緒にトランスフェクトした。空の対照であるpVax1プラスミドでトランスフェクトした細胞は、陰性対照として機能した。(C)EP介在送達後の抗CHIKV-IgG抗体のインビボ発現。マウス(B6.Cg-Foxn1
nu
/J)にCHIKV-IgGプラスミド(合計で100μg)を単回筋肉内注射で投与した後、EPを行った(1群当たりn=5マウス)。空のpVax1ベクターの注射は、陰性対照として用いた。
(D)CHIKV-Env抗原への特異的結合は、CHIKV-IgG及び組み換えCHIKV-Envで免疫化されたマウス由来の血清を回収したELISAアッセイにより測定し、異なる時点で個々のマウスのOD450nm値として表した。(E)ヒトIgG濃度の血清レベルは、実施例17に記載のように、CHIKV-IgGを筋肉内に注射したマウスにおいて種々の時点で測定した。(F)抗体結合親和性及び特異性の評価。タンパク質を標的にするためのCHIKV-IgGを注射したマウス(14日目)由来の血清の結合親和性機能性は、以下の実施例に記載のように、CHIKV感染細胞由来の細胞溶解物を用いてウエスタンブロットによって試験した。
CHIKV-IgG発現プラスミドの単回電気穿孔媒介注射後のIgGの発現及び結合速度を示す。(A)CHIKV-Fabを投与されたマウス由来の血清は、CHIKV-Env抗原に特異的であった。ELISAプレートを組み換えCHIKV-EnvまたはHIV-1 Env(亜型B;MN)タンパク質で被覆し、CHIKV-IgGまたはpVax1を注射したマウス由来の血清は、最初の注射後に示したように得られた。CHIKV-Env抗原への特異的結合は、血清を回収したELISAアッセイにより測定し、異なる時点で個々のマウスのOD450nm値として表した。(B)CHIKV-Fabを投与されたマウスから生成されたCHIKV-Fabが、CHIKV-Env糖タンパク質に結合できたことが、免疫蛍光検査法(IFA)結果により実証された。CHIKV感染ベロ細胞を感染後24時間で固定し、次いで免疫蛍光検査法でCHIKV-Env抗原発現を検出した。細胞核をDAPIで染色した。適量のCHIKV-Envタンパク質発現が、CHIKV-Fab抗体をもつベロ細胞中で観察された。pVax1免疫化マウス血清を陰性対照をとして用いた。(C)プラスミドを注射したマウス由来の血清の、CHIKV感染細胞への結合のFACS分析。x軸は、CHIKV-Envで補完したレンチウイルスGFP偽ウイルスを用いてGFP染色を示した。y軸は、マウス中で産生された試験ヒトIgGの染色を示した。二重陽性細胞は、CHIKV感染細胞への血清結合の示唆/測定であった。
CHIKV-IgG+EPを注射したマウス由来の血清が、多数のCHIKV株に対して中和活性を呈したことを示す。(A~F)6つの異なるCHIKVウイルス株:Ross、LR2006-OPY1、IND-63-WB1、PC-08、B448-China、及びBianchiに対して測定した、CHIKV-IgGをEPで投与したマウス由来の血清の中和活性を示す。中和抗体(nAb)力価は、ベロ細胞中でCPEの少なくとも50%阻害をもたらした血清の最大希釈としてプロットした。各実験において1群当たり少なくとも10匹のマウスである2つの独立した実験において、同様の結果が観察された。IC-50値は、Prism GraphPadソフトウェアで行った。
抗CHIKV-Env IgG及び血清の耐久性、及びCHIKV-Fabで免疫化した後の粘膜IgG応答、並びにIgG発現及び暴露試験を示す。(A)IgGプラスミド免疫化及びCHIKV暴露の概略図。(B~C)BALB/cマウスに、0日目にpVax1、CHIKV-IgG、またはCHIKV-Fabを注射し、2日目(B)または30日目(C)にCHIKV-Del-03(JN578247)CHIKV株(25ulの全容量中1×10
7
PFU)で暴露した。マウスを毎日監視し、生存率は、ウイルス暴露後20日間記録した。(D~E)異なる経路のCHIKVウイルス感染からのマウスの防御。2つの群のマウスは、筋肉内(IM)注射により100ugのCHIKV-IgGで免疫化し、皮下(s.c)(D)で2日目に暴露し(D)、別の群のマウスは、鼻腔内(i.n)により暴露した。(E)CHIKVを接種。マウスを毎日監視し、生存率は、ウイルス暴露後20日間記録した。↑は、DNA投与を示し;☆は、ウイルス暴露を示した。各群は、10匹のマウスからなり、結果は、2つの独立した実験の代表例であった。
抗CHIKV-Env IgG及び血清の耐久性、及びCHIKV-Fabで免疫化した後の粘膜IgG応答、並びにIgG発現及び暴露試験を示す。(A)IgGプラスミド免疫化及びCHIKV暴露の概略図。(B~C)BALB/cマウスに、0日目にpVax1、CHIKV-IgG、またはCHIKV-Fabを注射し、2日目(B)または30日目(C)にCHIKV-Del-03(JN578247)CHIKV株(25ulの全容量中1×10
7
PFU)で暴露した。マウスを毎日監視し、生存率は、ウイルス暴露後20日間記録した。(D~E)異なる経路のCHIKVウイルス感染からのマウスの防御。2つの群のマウスは、筋肉内(IM)注射により100ugのCHIKV-IgGで免疫化し、皮下(s.c)(D)で2日目に暴露し(D)、別の群のマウスは、鼻腔内(i.n)により暴露した。(E)CHIKVを接種。マウスを毎日監視し、生存率は、ウイルス暴露後20日間記録した。↑は、DNA投与を示し;☆は、ウイルス暴露を示した。各群は、10匹のマウスからなり、結果は、2つの独立した実験の代表例であった。
CHIKV暴露試験を介した即時的なな防御及び持続的な防御の双方を示す。(A)CHIKV-IgGワクチン接種及び暴露試験の概略図。I群暴露:BALB/cマウスに、0日目にCHIKV-IgG、CHIKV-Env、またはpVax1を注射し、2日目にCHIKV-Del-03(JN578247)ウイルス株(25ulの全容量中1×10
7
PFU)で暴露した。II群暴露:BALB/cマウスに、0日目に単回CHIKV-IgG免疫化または示した日に多数のCHIKV-Env免疫化のいずれかを施した後、I群暴露と同じ条件下で35日目に暴露した。↑は、DNA投与を示し;☆は、ウイルス暴露を示した。各試験において、マウスを20日間監視し、生存率を記録した。(B)I群暴露試験からのマウスの生存率曲線。100%生存率は、CHIKV-IgG免疫化マウスで記録された点に留意されたい。(C)II群暴露試験からのマウスの生存率曲線。
(D)抗CHIKVヒトIgGレベルの濃度は、CHIKV-IgG+EPで免疫化した後の示した時点で測定した。(E)マウスにおけるCHIKV-IgG及びCHIKV-Env免疫化した後の抗CHIKV-Env抗体の持続的かつ全身的な誘発。
CHIKVによる感染に応答したエクスビボサイトカイン産生を示す。(A)45日目(すなわち、暴露後10日目)のII群暴露試験からのCHIKV-IgG及びCHIKV-Env投与マウスにおけるウイルス力価。各データポイントは、10匹のマウスからの平均ウイルス力価を表した。pVax1免疫化マウスの群は、対照として機能した。ウイルス負荷は、pVax1マウスと比べてCHIKV-IgG(p=0.0244)及びCHIKV-Env(p=0.0221)の両方において有意に減少した。(B&C)CHIKV感染マウスからの血清炎症性サイトカインレベル(TNF-α及びIL-6)の特徴付け。サイトカインレベルは、特異的ELISAアッセイによって45日目(暴露後15日目)にマウスで測定した。CHIKV-IgGまたはCHIKV-Envを注射したマウスは、TNF-α及びIL-6の血清レベルが同様であり、かつ、対照群(p<0.0001)よりもTNF-α及びIL-6の血清レベルが有意に低かった。データは、マウス1匹当たり(1群当たりn=10)3つのウェルの平均を表した。
(D)CHIKV-IgGまたはCHIKV-Envで免疫化した後、CHIKV特異的ペプチドで刺激したマウスの脾細胞におけるT細胞応答。示したデータは、少なくとも2つの別々の実験の代表例であった。
抗体をコードするDNA構築物によって送達されたヒト抗DENV中和mAbのインビトロ発現をを示す。(a)送達に使用したDNAプラスミドの概略図;抗体重鎖及び軽鎖配列は、フリン及び2A切断部位の組み合わせによって分離する。(b)pDVSF-3 WTまたはLALAをトランスフェクトした293T細胞の上清におけるヒトIgGのELISA定量化分析。(c)DVSF-3 WTを含有する、pDVSF-3 WTをトランスフェクトした293T上清のウエスタンブロット分析。抗体をプロテインAスピンカラムで精製し、還元(左)及び非還元(右)条件下でSDS-PAGEで分離した。
(d)ベロ細胞を非感染(モック)かまたはDENV1、2、3、または4で感染させるかのいずれかの後、固定し、透過処理し、pDVSF-3 WTまたはLALAをトランスフェクトした293T細胞の上清で染色した。
マウス血清中の中和DENV抗体の長期発現の結果を示す。(a)ヒトIgGの全ての血清検出可能レベルは、抗DENVヒトIgG抗体DVSF-1をコードするDNAプラスミドをFoxn1/NuJ免疫不全マウスに単回筋肉内注射した後、ELISA法で測定した。0~4週目(左)及び19週目(右)のヒトIgGレベル。各線(左)または点(右)は、個々のマウスを表した(n=5)。(b)血清中の全ヒトIgGは、129/SvマウスにpDVSF-3 WTまたはpDVSF-3 LALAプラスミドを筋肉内注射した後にELISA法で測定した(1群当たりn=4~5)。
(c)ベロ細胞を非感染(モック)かまたはDENV1、2、3、または4で感染させるかのいずれかの後、固定し、透過処理し、pDVSF-3 WTまたはpDVSF-3 LALAのいずれかのDNA注射後0または7日目に採取した129/Svマウス血清で染色した(1群当たりn=5)。
(d)中和は、ベロ細胞に追加する前にpDVSF-3 WTまたはpDVSF-3 LALA(1群当たりn=5)のいずれかのDNA注射後7日目に採取した129/Svマウス血清の連続希釈物でDENV1、2、3、または4をインキュベートすることで評価した。感染細胞の割合を示す。
抗体をコードするDNA構築物の送達により、ウイルスのみ及び抗体感染増強疾患に対して防御したことを示す。(a)ウイルスのみ暴露:AG129マウスは、亜致死量のDENV2 S221に暴露する5日前にpDVSF-3 WT、pDVSF-3 LALA、またはpVax空ベクターのいずれかの筋肉内注射を受けた(1群当たりn=5~6;pDVSF-3 LALA及びpDVSF-3 WTの比較に対するp≦0.0084)。(b)抗体依存性感染増強暴露:AG129マウスは、高用量の非中和抗DENVmAb 2H2を投与する5日前にpDVSF-3 WT、pDVSF-3 LALA、またはpVax空ベクターのいずれかの筋肉内注射を受けた。30分後、マウスに亜致死量のDENV2 S221を暴露した(1群当たりn=5~6;pDVSF-3 LALA及びpDVSF-3 WTの比較に対するp≦0.0072)。カプランマイヤー法による生存率曲線を(a~b)に示す。
pDVSF-3 WT及びLALAコード抗体のインビトロ機能分析を示す。(a)精製した組み換えDENV Eタンパク質に対する、pDVSF-3 WTまたはLALAをトランスフェクトした293T細胞の上清中のヒトIgGのELISA結合分析。(b)抗体依存性感染増強は、K562細胞に追加する前にpDVSF-3 WTまたはLALAの上清の連続希釈物でDENV1、2、3、または4をインキュベートすることで評価した。感染細胞の割合を示す。
抗体をコードするDNA構築物を送達した後のAG129マウス中の抗DENVヒトIgGレベルの暴露前レベルを示す。(a)血清中のDVSF-3 WTまたはDVSF-3 LALAの全ヒトIgGは、AG129マウス中のそれぞれのプラスミドをDNA筋肉内注射(DENV2暴露1日前)及びEPした4日後にELISA法で測定した(1群当たりn=5~6;pDVSF-3 WT及びpVaxの比較に対するp≦0.0005;pDVSF-3 LALA及びpVaxの比較に対するp≦0.0001)。
増加したDENV1-4抗血清を産生したマウスにおける多数のDENV抗体コードプラスミドの送達を示す。(a)血清中のDVSF-3 WT、DVSF-1 WT、またはDVSF-3 WT、及びDVSF-1 WTの全ヒトIgGは、129/Svマウス中のそれぞれのプラスミドをDNA筋肉内注射及びEPした7日後にELISA法で測定した(1群当たりn=5;pDVSF-1 WT及びpDVSF-1+3の比較に対するp≦0.0088;pDVSF-3 WT及びpDVSF-1+3の比較に対するp≦0.0240)。
上部パネルには、DVSF-3 WTは、ヒトFcyR1aに結合したが、DVSF-3 LALAは、FcyR1aに結合しなかったことを示す。下部の4つのパネルは、抗体依存性感染増強アッセイの結果を示す:DVSF-3 LALAによるDENV-1、-2、-3、または-4のインキュベーションは、ヒト単球(K562細胞株)感染をもたらさなかったが、DVSF-3 WTは、DENV-1、-2、及び-3の感染を高めた。
【発明を実施するための形態】
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
個人
DNA増幅システム
2か月前
東洋紡株式会社
逆転写反応用組成物
19日前
三菱製紙株式会社
パルプの糖化方法
2か月前
株式会社ブラスト
細胞培養装置
1か月前
学校法人近畿大学
高感度PCR法
2か月前
株式会社ブラスト
密閉チャンバー
1か月前
澁谷工業株式会社
液体吸引装置
2か月前
国立大学法人 琉球大学
多糖の製造方法
3か月前
国立大学法人 東京大学
ゲノム編集技術
1か月前
相生ユニビオ株式会社
ウイスキーの製造方法
25日前
学校法人立命館
紐状構造物の製造方法
18日前
小林製薬株式会社
黒ずみ形成方法
1か月前
個人
急性ストレス評価用データの生成方法
5日前
国立大学法人神戸大学
油脂生産方法
3か月前
東洋紡株式会社
核酸増幅用組成物の保存安定性向上方法
3か月前
松谷化学工業株式会社
アルコール飲料及びその製造方法
1か月前
株式会社パウレック
培養装置及び培養方法
1か月前
サッポロビール株式会社
ビールテイスト飲料
1か月前
株式会社東海ヒット
完全閉鎖型灌流液送液系
25日前
学校法人明治大学
キラーT細胞の活性化方法
3か月前
国立大学法人東京海洋大学
熱処理リゾチーム組成物
24日前
花王株式会社
リパーゼ変異体
1か月前
株式会社デンソー
バイオセンサ装置
1か月前
日本精工株式会社
分注装置
2か月前
花王株式会社
変異プロテアーゼ
3か月前
個人
脱炭酸システム、ハイドロゲルおよびその製造方法
3か月前
サッポロビール株式会社
ビールテイスト飲料
1か月前
横河電機株式会社
核酸抽出方法
5日前
サッポロビール株式会社
ビールテイスト飲料
1か月前
サッポロビール株式会社
ビールテイスト飲料
1か月前
サッポロビール株式会社
ビールテイスト飲料
1か月前
サッポロビール株式会社
ビールテイスト飲料
1か月前
サッポロビール株式会社
ビールテイスト飲料
1か月前
株式会社ヤマザキエンジニアリング
製麹装置及び製麹方法
1か月前
花王株式会社
リパーゼの探索方法
2か月前
NTN株式会社
細胞組織の製造方法
1か月前
続きを見る
他の特許を見る








 特許ウォッチ
特許ウォッチ