TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025089506
公報種別
公開特許公報(A)
公開日
2025-06-12
出願番号
2025052481,2022577265
出願日
2025-03-26,2020-07-14
発明の名称
γ-グロビン遺伝子発現を活性化する方法、および組成物
出願人
広州瑞風生物科技有限公司
,
GUANGZHOU REFORGENE MEDICINE CO.,LTD.
代理人
弁理士法人太陽国際特許事務所
主分類
C12N
15/113 20100101AFI20250605BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】本発明は、γ-グロビン遺伝子転写を活性化する新方法を開示する。
【手段】本発明は、GATAまたはそのアンチセンス相補配列TATCを含有する一本鎖オリゴヌクレオチド(ssODN)をガイド情報として用い、γ-グロビン遺伝子調節領域において遺伝子編集を行ってGATAを含有するエンハンサー要素を形成することにより、γ-グロビン遺伝子の成熟した赤血球における発現を促進することができる。本発明に係る技術を利用して遺伝子編集した造血幹細胞は、正常な機能を有し、且つ赤血球に分化した後、胎児ヘモグロビンの発現を顕著に向上させることができるため、β-地中海貧血症および鎌型赤血球貧血症の臨床治療に用いることができる。
【選択図】図1
特許請求の範囲
【請求項1】
γ-グロビン遺伝子発現を活性化する方法であって、
遺伝子編集技術によってγ-グロビン遺伝子調節領域におけるセンス鎖またはアンチセンス鎖に、GATAまたはTATC配列を人為的に導入して、γ-グロビン遺伝子の調節領域においてGATAモチーフを含むエンハンサー要素を形成することを含む、方法。
続きを表示(約 830 文字)
【請求項2】
GATAまたはTATC配列の導入は、配列の削除、配列の挿入、配列の変異、またはそれらの組合せにより実現される、
請求項1に記載の方法。
【請求項3】
GATAモチーフは、WGATAR配列、または対応するアンチセンス相補配列YTATCWであり、ただし、Wは、TまたはAであり、Rは、AまたはGであり、Yは、TまたはCである、
請求項1に記載の方法。
【請求項4】
前記エンハンサー要素には、NTGモチーフまたはNAGモチーフ、または対応するアンチセンス相補配列がさらに含まれ、
ただし、Nは、A、G、CまたはTである、
請求項1に記載の方法。
【請求項5】
前記GATAモチーフとNTGモチーフまたはNAGモチーフとの距離が7~8塩基であり、且つ前記NTGモチーフまたはNAGモチーフが前記GATAモチーフの上流に位置する、
請求項4に記載の方法。
【請求項6】
GATAまたはTATC配列が含まれる、遺伝子編集用ssODN。
【請求項7】
前記ssODNには、5′相同アーム、置換配列、3′相同アームが含まれる、
請求項6に記載のssODN。
【請求項8】
前記ssODNにおける5′相同アームと3′相同アームとが対称的または非対称的であり、5′相同アームと3′相同アームの長さが20~300ntである、
請求項7に記載のssODN。
【請求項9】
前記ssODNにおける5′相同アームまたは3′相同アームは、擬似編集領域DNAのセンス鎖またはアンチセンス鎖から選ばれる、
請求項7に記載のssODN。
【請求項10】
前記ssODNにおける置換配列の塩基の数が0~6である、
請求項7に記載のssODN。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、遺伝子編集の技術分野に関し、γ-グロビン遺伝子発現を活性化するための方法、組成物およびその使用に関する。当該方法は、GATAまたはそのアンチセンス相補配列TATCを含有する一本鎖オリゴヌクレオチド(ssODN)をガイド情報として用い、γ-グロビン遺伝子のプロモーター領域において遺伝子編集を行い、GATAを含有する赤血球系エンハンサー要素を形成することにより、γ-グロビン遺伝子の成熟した赤血球における発現を促進することができる。
続きを表示(約 4,700 文字)
【背景技術】
【0002】
ヘモグロビン(hemoglobin、単にHbと呼ばれる)は、赤血球内で酸素ガスを携帯し搬送するための特殊なタンパク質である。ヘモグロビンは、グロビン(globin)およびヘムからなる。成人の体内のヘモグロビンは、主に2つのα-グロビンおよび2つのβ-グロビンからなるテトラマー(α2β2)であり、成人ヘモグロビン(HbA)と呼ばれる。β-グロビンの遺伝子(HBB)の変異は、鎌状赤血球症(Sickle Cell Disease:SCD)およびβ-地中海貧血症(β-thalassemia、単にβ-thalと呼ばれる)を含むβ-ヘモグロビン障害(β-ヘモグロビン症とも呼ばれる)を引き起こす可能性がある。鎌型赤血球貧血症は、β-グロビン構造遺伝子の点変異により引き起こされるものであり、異常のヘモグロビン(HbS)を発生することがある。β-地中海貧血症は、β-グロビン遺伝子発現の一部または全部の欠陥により引き起こされるものであり、成人ヘモグロビン(HbA)の欠陥または欠損を引き起こすことがある。ヘモグロビンのグロビン鎖の減少または欠損は、ヘモグロビンの構造が異常となることを引き起こす可能性がある。このような異常のヘモグロビンを含有する赤血球は、変形性が低下し、寿命が短く、骨髄中にインサイツ溶血が発生する可能性があり、末梢血液循環に進入した後、脾臓などの臓器により繰り上げて破壊され、貧血、体内の鉄沈着、ひいては発育異常を引き起こす可能性がある。ヘモグロビン症は、世界の数百万の人口に影響を及ぼしており、現在、毎年約330,000の子供がヘモグロビン症で生まれ、人類の健康や生命を深刻に脅かしている。地中海貧血症および鎌状赤血球症の患者は、主に、規範的な長期輸血と脱鉄治療によって病状を緩和するが、このような形態によって病状が治癒できないだけでなく、高いセキュリティリスクが存在している。異体造血幹細胞移植は、現在、地中海貧血症および鎌状赤血球症を根治できる唯一の治療技術であるが、骨髄マッチング成功率が低く、および免疫拒絶リスクが存在するなどの要素の制限により、臨床では、広く適用することが困難である。その為、新たな安全的で有効な治療方法を開発する必要性が大きくなっている。
【0003】
ヒトの発育過程では、ヘモグロビンは、常に、2つのα-グロビンおよび2つのβ-グロビンの形態で構成されているわけではない。胚の発育期間および出生直後、ヘモグロビンは、2つのα-グロビン鎖および2つのγ-グロビン鎖からなるテトラマーの形態(α2γ2)で存在し、胎児ヘモグロビン(HbF)と呼ばれ、HbAに対してより強い酸素親和力を持っている。胎児の発育に伴い、γ-グロビン遺伝子がだんだん発現せずに沈黙するが、同じゲノムサイト付近のβ-グロビン遺伝子の発現がだんだん上昇し、赤ちゃんの生後約半年以降、血液におけるヘモグロビンの成分および割合がだんだん安定し、HbFが成人ヘモグロビン(HbA)で置換され、極めて低いレベルのHbFだけが残されている(総ヘモグロビンの1%以下を占める)。
【0004】
検討から分かるように、一部の人たちのうち、ある特殊なサイトに変異が現れ、γ-グロビン遺伝子転写が活性化されて、ヘモグロビンにおけるHbFの割合が向上してしまう(遺伝性高胎児ヘモグロビン血症:hereditary persistence of fetal hemoglobin、HPFHとも呼ばれる)。たとえば、γ-グロビン遺伝子のプロモーター領域において幾つかの異なる種類の変異、たとえば-114~-102 13bp del c .、4bp del c .-225~-222、c .-114C>T、c .-117G>A、c .-158C>T、c .-167C>T、c .-170G>A、c .-175T>G、c .-175T>C、c .-195C>G、c .-196C>T、c .-198T>C、c .-201C>Tなどが発見されている。これらの自然に存在する変異は、異なる程度でHbFの総ヘモグロビンに対する割合を向上させることができる。臨床検討から分かるように、少数のβ-地中海貧血症または鎌型赤血球貧血症の患者は、ゲノムにHPFH変異がちょうどよく存在するため、HbFの発現がHbAの不足を補い、ある程度で患者の貧血病状を軽減または緩和したり、輸血のニーズを低減したりすることができる。この発見に啓発されると、科学者らは、β-ヘモグロビン症を治療する目的を達成するために、継続して各種のHbF発現を誘導可能な方法を探索している。たとえば、現在、臨床ではヒドロキシ尿素などの薬物を用いてHbF発現を誘導してβ-ヘモグロビン症を治療することがあるが、数多くの患者に対してHbFの誘導レベルが低く、患者の状況を徹底的に改善することができない。
【0005】
遺伝子編集技術は、ヘモグロビン症などの遺伝病の治療に対して新希望および新方法をもたらしている。近年、遺伝子編集技術は、画期的な開発し、ゲノムにおける特定のサイトの塩基配列を人為的に変更することが可能となるとともに、ますます容易になる。現在、発展が相対的に成熟した遺伝子編集技術は、ZFN(ジンクフィンガーヌクレアーゼ)、TALEN(転写活性因子様エフェクターヌクレアーゼ)およびCRISPR(クラスター化され、規則的に間隔が空いている短い回文の反復)/Casシステム(CRISPR-Casシステム)である。特異の遺伝子編集システムを設計し、生細胞における目標ゲノムDNAサイトを切断して二本鎖切断(DSB)を形成することにより、細胞が非相同末端結合(non-homologous end joining:NHEJ)修復仕組みを利用して、DNA欠陥を修復し、配列の挿入、欠損、塩基置換などの変異をランダム形成することができる。たとえば、遺伝子編集を行うと同時に、人為的に設計されたDNAドナーテンプレート(donor template)を提供することにより、細胞が相同組換え修復(homology-directed repair:HDR)仕組みを利用して修復を完成することで、目標サイトに所望する塩基変異形態を導入することができる。
【0006】
現在、遺伝子編集によって赤血球におけるHbF発現を補強する治療策略としては、国際的に幾つかの異なる方法がある。(1)ヒト2番染色体におけるBCL11A遺伝子のイントロンにおける赤血球系エンハンサー要素を削除または破壊し、赤血球におけるBCL11Aの発現を低減させ、BCL11Aのγ-グロビン遺伝子転写に対する抑制作用を解除する。(2)γ-グロビン遺伝子のプロモーター領域における一部の配列を削除または破壊し、転写抑制因子の結合を阻止したり、自然に存在するHPFH変異種類、たとえば-114~-102サイトの13塩基対の削除(13bp del c.)を導入したりする。(3)3.5kb~13.6kbの大きなセグメントのDNA削除を製造し、γ-グロビン遺伝子とβ-グロビンとの間の不明の抑制因子を含有する配列を削除して、γ-グロビン遺伝子が遠端LCRエンハンサーと結合することを促進する。これらの方法は、HbF発現を活性化する効果を示しているが、その長期間効果または/および安全性がまだ明確ではなく、新たな方法をさらに開発する必要がある。
【発明の概要】
【発明が解決しようとする課題】
【0007】
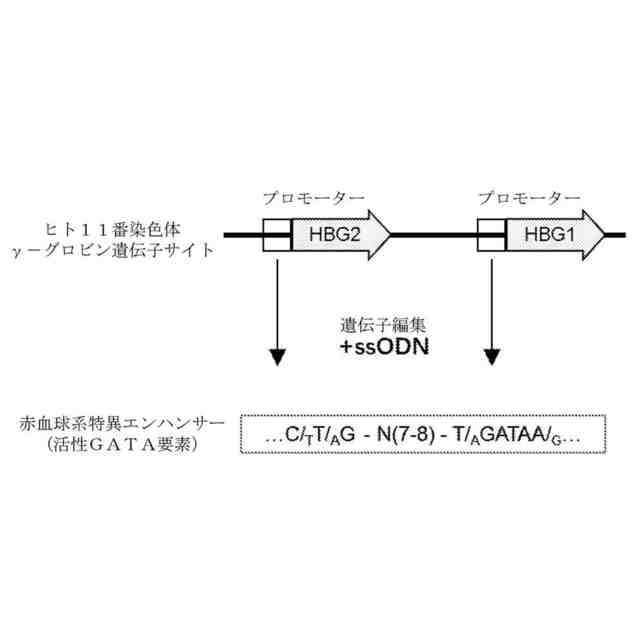
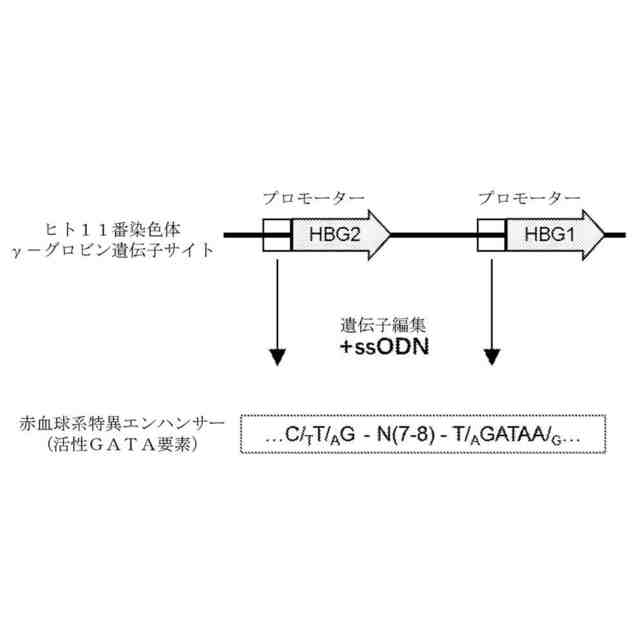
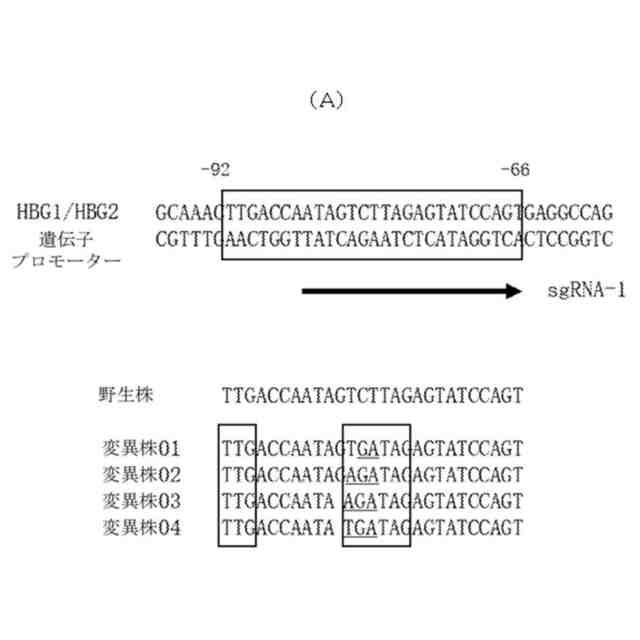
本発明は、遺伝子編集によってγ-グロビン遺伝子転写を活性化する方法を提供する。本発明は、GATAまたはそのアンチセンス相補配列TATCを含有する一本鎖オリゴヌクレオチド(ssODN)を利用して、遺伝子編集技術(例えば、CRISPR-Cas遺伝子編集システム)によって、γ-グロビン遺伝子のプロモーター領域における適当な箇所に少数の塩基の変異(削除、置換、挿入など)を製造し、編集した後にγ-グロビン遺伝子のプロモーター領域のセンス鎖またはアンチセンス鎖にGATAを含有する赤血球系エンハンサー要素、例えば、NTG-N(7-8)-WGATARまたはNAG-N(7-8)-WGATARの配列構造を形成することで、γ-グロビン遺伝子の成熟した赤血球における転写を促進し、赤血球におけるHbF発現を向上させることができる。そのうち、Wは、TまたはAであり、Rは、AまたはGであり、Nは、A、G、CまたはTであり、CまたはTであることがより好ましい。N(7-8)とは、7~8個の任意の塩基を指す。
【0008】
図1に示すように、本発明の原理は、CRISPR-Casなどの遺伝子編集技術に基づき、本発明に係る一本鎖オリゴヌクレオチド(ssODN)を結び付けて、CD34+造血幹細胞などの赤血球分化能力を有する幹細胞、前駆細胞に対して効率的な遺伝子編集を行い、コードされたヒトγ-グロビンのHBG1及びHBG2遺伝子(ヒト11番染色体に位置する)プロモーターにおいて変異を製造し、プロモーターのセンス鎖またはアンチセンス鎖において1つのNTG-N(7-8)-WGATARまたはNAG-N(7-8)-WGATARの配列構造を人為的に創造する。当該配列は、エンハンサーとして、目的細胞が赤血球に分化した後にGATA1などの活性化因子を募集してγ-グロビン発現を促進することである。
【0009】
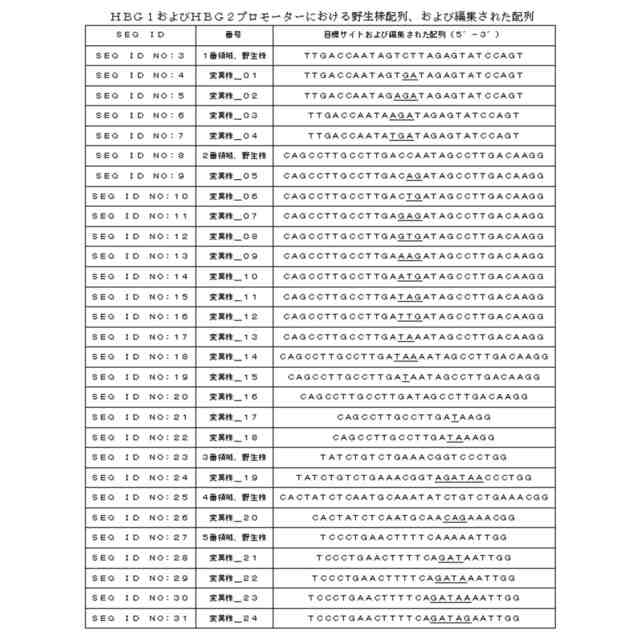
本発明は、GATAまたはTATC配列を含有するssODNを提供する。当該ssODNは、γ-グロビン遺伝子の遺伝子編集に用いることができる。本発明では、ssODN構造には、5′相同アーム、置換配列、3′相同アームが含まれる。そのうち、GATAまたはTATC配列は、ssODNにおける任意の箇所、たとえば置換配列、5′相同アーム、3′相同アーム、及び5′相同アームと置換配列との結合箇所、3′相同アームと置換配列との結合箇所に位置してもよい。片側の相同アームの長さは、20~300ntであってもよい。本発明の実施例において、ssODN相同アームの長さは、40nt程度である。本発明の一部の実施例において、GATAまたはTATC配列が含まれたssODNは、SEQ ID NO:40~SEQ ID NO:65で表される配列から選ばれる。そのうちの一部の実施例において、ssODNの5′および3′は、チオリン酸(Phosphorothioate)で修飾されたものである。
【0010】
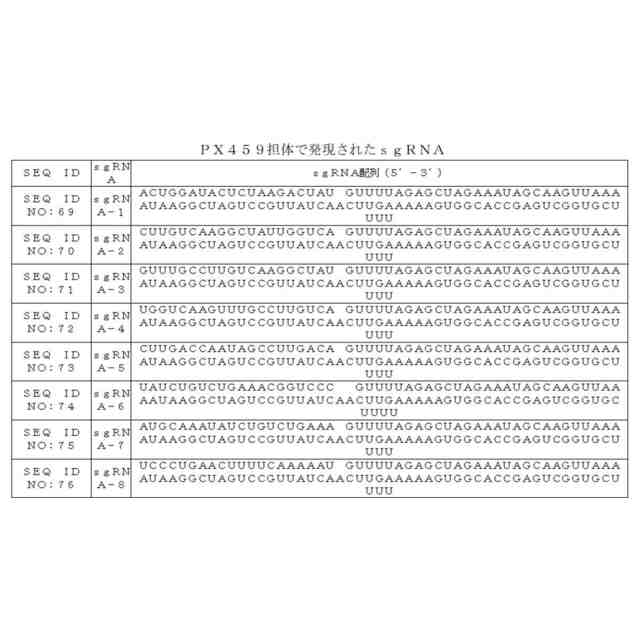
本発明は、γ-グロビン遺伝子編集用組成物を開示する。当該組成物には、ssODN、sgRNA、およびCRISPR-Casヌクレアーゼが含まれる。本発明の一部の実施例において、前記遺伝子編集システムにおけるsgRNA標的サイトがSEQ ID NO:32~SEQ ID NO:39で表される配列に位置する。本発明の一部の実施例において、前記sgRNAは、SEQ ID NO:69~SEQ ID NO:76で表される配列である。本発明の一部の実施例において、前記Cas9タンパク質は、Streptococcus pyogenesに由来するCas9タンパク質である。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
合同酒精株式会社
麦汁の製造方法
2か月前
池田食研株式会社
RNAの合成方法
2か月前
杏林製薬株式会社
核酸検出用PCR溶液
17日前
サッポロビール株式会社
飲料
13日前
学校法人近畿大学
培養肉の製造方法
1か月前
JNC株式会社
アデノ随伴ウイルスの精製方法
1か月前
朝日酒造 株式会社
発泡性清酒の製造方法
2か月前
セージ セラピューティクス, インコーポレイテッド
C7、C12、およびC16置換神経刺激性ステロイドおよびそれらの使用方法
2か月前
株式会社東海ヒット
灌流培養ユニット
4日前
個人
ナノ微粒子の製造方法
13日前
ヤマト科学株式会社
インキュベータ
1か月前
個人
超音波機能着きウィスキー熟成ボトル用のキャップ
28日前
日本特殊陶業株式会社
メタン発生抑制装置
1か月前
大和ハウス工業株式会社
藻類培養システム
2か月前
株式会社アテクト
培養シート
1か月前
東洋紡株式会社
緩衝剤によるヘムタンパク質の安定化方法
28日前
住友化学株式会社
細胞チップ
2か月前
学校法人慈恵大学
培養皿用の定規
2か月前
花王株式会社
肌タイプの分類方法
1か月前
学校法人藤田学園
アンチセンス核酸およびその利用
20日前
個人
移植材料の評価方法
今日
キヤノン株式会社
細胞剥離装置
2か月前
ニプロ株式会社
コネクタ及びバッグ
1か月前
学校法人東京電機大学
酵母、及び食品用添加剤
5日前
学校法人上智学院
生物の管理方法
2か月前
学校法人上智学院
核酸塩基検出剤
27日前
学校法人近畿大学
遺伝子組換えコクナーゼおよびその前駆体
1か月前
カルビー株式会社
睡眠の質を評価する方法
11日前
住友ベークライト株式会社
細胞培養キット
2か月前
関西電力株式会社
微細藻類培養システム及び微細藻類培養方法
1か月前
国立大学法人東京科学大学
慢性腎臓病の検査方法
2か月前
横河電機株式会社
伝熱装置、加熱装置及び処理装置
1か月前
国立大学法人 新潟大学
試料製造方法
26日前
花王株式会社
体脂肪変化の予測方法
13日前
ネクスジェン株式会社
細胞の培養方法および組成物
27日前
キッコーマン株式会社
甘味が増強された酒類
13日前
続きを見る
他の特許を見る




 特許ウォッチ
特許ウォッチ