TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025084823
公報種別
公開特許公報(A)
公開日
2025-06-03
出願番号
2025027824,2022551605
出願日
2025-02-25,2021-03-01
発明の名称
リプログラミングされたtracrRNAを用いたRNA検出及び転写依存性編集
出願人
ヘルムホルツ-ツェントルム フュア インフェクツィオンスフォルシュンク ゲーエムベーハー
,
ユリウス-マクシミリアンズ-ウニヴェルジテート ウュルツブルグ
代理人
個人
主分類
C12N
15/11 20060101AFI20250527BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】特定のRNAを検出するための方法を提供する。
【解決手段】本発明は、細胞、組織、及び/又は試料中の少なくとも1つの被検知RNAを、上記被検知RNAと特異的にハイブリダイズする少なくとも1つの非天然産生型tracrRNA、及び少なくとも1つの標的核酸に結合する少なくとも1つのtracrRNA依存性CRISPRヌクレアーゼ酵素を使用して検出する方法、並びにそれぞれのシステム、並びにその診断的使用及び治療的使用に関する。
【選択図】図1-1
特許請求の範囲
【請求項1】
天然のcrRNA:tracrRNA二重鎖を模倣する複合体の形成を通じて予め選択された被検知RNA配列に特異的にハイブリダイズするように設計されているアンチリピート領域配列を含む部分を含む、非天然産生型tracrRNA核酸分子。
続きを表示(約 3,100 文字)
【請求項2】
請求項1に記載の非天然産生型tracrRNA核酸分子と、少なくとも1つのtracrRNA依存性CRISPRヌクレアーゼ酵素と、少なくとも1つの被検知RNAとを含む複合体。
【請求項3】
少なくとも1つの被検知RNA配列に基づく配列、又は該被検知RNA配列に基づいて設計された配列を含む標的DNA核酸分子に更に結合される、請求項2に記載の非天然産生型tracrRNA核酸分子。
【請求項4】
細胞、組織、及び/又は試料中の少なくとも1つの被検知RNAを検出する方法であって、
a)前記試料と、前記被検知RNAの第1の部分と特異的にハイブリダイズする部分を含む少なくとも1つの非天然産生型tracrRNAとを接触させることと、
ここで、前記被検知RNAの第2の部分は、少なくとも1つの標的核酸と特異的にハイブリダイズし、前記少なくとも1つの非天然産生型tracrRNAは、任意にRNase III等の少なくとも1つのRNA切断酵素の存在を更に含む少なくとも1つのtracrRNA依存性CRISPRヌクレアーゼ酵素の存在下で前記被検知RNAとハイブリダイズするか、又はハイブリダイズすることができ、
b)前記少なくとも1つのtracrRNA依存性CRISPRヌクレアーゼ酵素の前記少なくとも1つの標的核酸への結合を検出することと、
を含み、
c)ここで、結合により、前記細胞、組織、及び/又は試料中の前記少なくとも1つの被検知RNAが検出され、かつ、
好ましくは、前記ヌクレアーゼは、その切断活性及び/又はニッキング活性が不活性化されている、方法。
【請求項5】
細胞及び/又は試料中の少なくとも1つの被検知RNAを検出する方法であって、
a)前記試料と、前記被検知RNAの第1の部分と特異的にハイブリダイズする部分を含む少なくとも1つの非天然産生型tracrRNAとを接触させることと、
ここで、前記被検知RNAの第2の部分は、少なくとも1つの標的核酸と特異的にハイブリダイズし、前記少なくとも1つの非天然産生型tracrRNAは、任意にRNase III等の少なくとも1つのRNA切断酵素の存在を更に含む少なくとも1つのtracrRNA依存性CRISPRヌクレアーゼ酵素の存在下で前記被検知RNAとハイブリダイズするか、又はハイブリダイズすることができ、
b)前記試料中の前記少なくとも1つの標的核酸の前記ヌクレアーゼ酵素による切断を検出することと、
を含み、
c)ここで、少なくとも1つの標的核酸の前記切断の検出により、前記細胞、組織、及び/又は試料中の前記少なくとも1つの被検知RNAが検出される、方法。
【請求項6】
前記結合された及び/又は切断された少なくとも1つの標的核酸の検出は、色素、蛍光体等の適切な標識のシグナル、又は導電率における変化を検出すること、及び/又は前記切断された少なくとも1つの標的核酸断片自体を検出することを含む、請求項4又は5に記載の方法。
【請求項7】
細胞、組織、及び/又は試料中の少なくとも1つの被検知RNAを検出する方法であって、
a)前記試料と、前記被検知RNAの第1の部分と特異的にハイブリダイズする部分を含む少なくとも1つの非天然産生型tracrRNAとを接触させることと、
ここで、前記被検知RNAの第2の部分は、少なくとも1つの標的核酸と特異的にハイブリダイズし、前記少なくとも1つの非天然産生型tracrRNAは、任意にRNase III等の少なくとも1つのRNA切断酵素の存在を更に含む少なくとも1つのtracrRNA依存性CRISPRヌクレアーゼ酵素の存在下で前記被検知RNAとハイブリダイズするか、又はハイブリダイズすることができ、
b)前記試料中の前記少なくとも1つの標的核酸を前記ヌクレアーゼによりニッキング又は切断することと、
c)例えば、非相同末端結合(NHEJ)修復、マイクロホモロジー媒介末端結合(MMEJ)、相同配列依存的修復(HDR)、塩基編集、プライム編集、又はRNA編集を含む、前記少なくとも1つの標的核酸を検出可能に編集することと、
d)例えば、インデル又は遺伝子編集の存在及び/又は長さ、前記少なくとも1つの標的核酸によってコードされる遺伝子又は他の遺伝要素の活性化/抑制の検出、及び/又は前記プライム編集又は塩基編集の検出から選択される少なくとも1つの方法を含む、前記少なくとも1つの標的核酸の前記編集を適切に検出することと、
を含み、
e)ここで、前記少なくとも1つの標的核酸の前記編集の検出により、前記細胞、組織、及び/又は試料中の前記少なくとも1つの被検知RNAが検出される、方法。
【請求項8】
細胞、組織、及び/又は試料中の少なくとも1つの標的DNAの転写を記録する方法であって、
a)前記試料、細胞、又は組織と、被検知RNAの第1の部分と特異的にハイブリダイズする部分を含む少なくとも1つの非天然産生型tracrRNAとを接触させることと、
ここで、前記被検知RNAの第2の部分は、前記少なくとも1つの標的DNAと特異的にハイブリダイズし、前記少なくとも1つの非天然産生型tracrRNAは、任意にRNase III等の少なくとも1つのRNA切断酵素の存在を更に含む少なくとも1つのtracrRNA依存性CRISPRヌクレアーゼ酵素の存在下で前記被検知RNAとハイブリダイズするか、又はハイブリダイズすることができ、
b)前記試料中の前記少なくとも1つの標的核酸を前記ヌクレアーゼによりニッキング又は切断することと、
c)非相同末端結合(NHEJ)修復、マイクロホモロジー媒介末端結合(MMEJ)、相同配列依存的修復(HDR)、検出可能なマーカー、検出可能な改変、塩基編集、プライム編集、又はRNA編集を含む、前記少なくとも1つの標的DNAを検出可能に編集することと、
d)任意に、インデル又は遺伝子編集の存在及び/又は長さ、前記少なくとも1つの標的核酸によってコードされる遺伝子又は他の遺伝要素の活性化/抑制の検出、及び/又は前記プライム編集又は塩基編集の検出から選択される少なくとも1つの方法を含む、前記少なくとも1つの標的DNAの前記編集を適切に検出することと、
を含み、
e)ここで、前記少なくとも1つの標的DNAの前記編集の検出により、前記細胞、組織、及び/又は試料中の前記少なくとも1つの被検知RNAの転写が記録される、方法。
【請求項9】
1つ以上の試料、組織、及び/又は細胞において、幾つかの又は多数の被検知RNAが検出される、請求項4~8のいずれか一項に記載の方法。
【請求項10】
前記少なくとも1つの被検知RNAは、環境、種、株、疾患、細胞、及び/又は組織に特異的であるか、又はウイルス感染症、コロナウイルス感染症、病原体による感染症、代謝疾患、癌、神経変性疾患、加齢、薬物、及び生物的ストレス又は非生物的ストレスから選択される病態に関連する、請求項4~9のいずれか一項に記載の方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、細胞、組織、及び/又は試料中の少なくとも1つの被検知RNAを、上記被検知RNAと特異的にハイブリダイズする少なくとも1つの非天然産生型tracrRNA、及び少なくとも1つの標的核酸に結合する少なくとも1つのtracrRNA依存性CRISPRヌクレアーゼ酵素を使用して検出する方法、並びにそれぞれのシステム、並びにその診断的使用及び治療的使用に関する。
続きを表示(約 4,500 文字)
【背景技術】
【0002】
殆ど全ての古細菌及び約半分の細菌は、クラスターを形成し規則正しい間隔を持つ短いパリンドロームリピート(CRISPR)-CRISPR関連遺伝子(Cas)適応免疫システムを有しており、これにより、原核生物はウイルス及び核酸ゲノムを有する他の外来侵入者から保護されている。CRISPR-Casシステムは、エフェクター複合体の組成に従ってクラス1とクラス2とに機能的に分類される。クラス2はシングルエフェクターヌクレアーゼからなり、II型、V型、及びVI型のCRISPR-Casシステムを含むクラス2のCRISPR-Casシステムの活用によって、ゲノム編集の慣行が実現されている。II型及びV型は、DNA又はRNAの標的化に使用され得るのに対して、VI型は、RNAの標的化に使用される(例えば、非特許文献1を参照)。II型及びV型のCasエフェクターヌクレアーゼは、通常、標的DNA認識における最初の工程としてプロトスペーサー隣接モチーフ(PAM)に依存し、エフェクターヌクレアーゼは、タンパク質-DNA相互作用を介してPAM配列に直接的に結合した後に、下流のDNA配列をアンジッピングする。次に、エフェクタータンパク質は、DNA標的の一方の鎖とCRISPR RNA(crRNA)のガイド部分との間の塩基対合の程度を照合する。2つの間の相補性が十分であると、標的の切断が駆動される。PAM配列は、システム間だけでなくそれ以外は類似のヌクレアーゼ間でもかなり異なることが知られており、Casタンパク質を操作してPAM認識を変化させることができることが示された(非特許文献2)。DNAの標的化に加えて、C.ジェジュニ(C. jejuni)のCas9、N.メニンギティディス(N. meningitidis)のCas9、S.アウレウス(S. aureus)のCas9、及び難培養古細菌由来のCas12f1等の一部のII型及びV型のシングルエフェクターヌクレアーゼは、ssDNA及び/又はRNAを標的化することも示されている(非特許文献3、非特許文献4、非特許文献5)。これらの場合に、PAMは必要とされなかった。S.ピオゲネス(S. pyogenes)のCas9(SpyCas9)等の一部のヌクレアーゼは、ssDNA又はRNAを直ちに標的化することができなかったが、オリゴヌクレオチドを供給して二本鎖PAM領域を生成することで、SpyCas9が一本鎖標的に結合してそれを切断することができた(非特許文献6)。
【0003】
成熟crRNAは、侵入ゲノムに対するCRISPR-Cas防御における重要な要素である。これらの短いRNAは、同種の侵入核酸にCasタンパク質(複数の場合もある)を導いてそれらを破壊する特有のガイド配列を含む。CRISPR RNA(crRNA)は、保存された交互のリピート及びスペーサーを含むCRISPRアレイにおいて天然にコードされている。このアレイは、長いpre-crRNAに転写されることが多く、次にこのRNAは、リピート-スペーサー単位の部分から構成される個々のcrRNAへとプロセシングされる。様々な種類のCRISPR-Casシステムは、別個のcrRNA成熟メカニズムを進化させてきた。多数のサブタイプ(II-A、II-B、II-C、V-B、V-C等)は、独自のcrRNA生合成経路を進化させており、そこでは、トランス活性化型CRISPR RNA(CRISPR-Cas遺伝子座内にコードされたtracrRNA)がpre-crRNAの各リピート配列と塩基対合して、二本鎖のRNA二重鎖を形成する(非特許文献7)。crRNA-tracrRNA二重鎖は、ハウスキーピングエンドリボヌクレアーゼRNase IIIによって切断され、システムに特異的なシングルエフェクターヌクレアーゼが結合する。次に、このヌクレアーゼは、RNA二重鎖内のcrRNAを使用して、DNA標的化、場合によってはRNA標的化に導く。これまでに、Cas9、Cas12b1/C2c1、Cas12b2、Cas12e/CasX、Cas12f1/Cas14a、Cas12g、及びCas12kを含む多数のtracrRNA依存性CRISPRヌクレアーゼが報告されている。一連のトランスポゾン遺伝子と連携して機能してDNAテンプレートのRNA指令型挿入を行うため、Cas12kヌクレアーゼは独特である(非特許文献8)。これらの各場合において、tracrRNA:crRNA二重鎖は、シングルガイドRNA(sgRNA)と呼ばれる単一のRNAキメラとして操作された場合に、Cas9及びその他のtracrRNA依存性CRISPRヌクレアーゼによる配列特異的なdsDNA切断も誘導する(非特許文献9、非特許文献10を参照)。
【0004】
フランシセラ・ノビシダ(Francisella novicida)のCRISPR-Cas9システムは、Cas9とその内因性DNA標的とのPAM依存性相互作用を示し、これは、非正規の低分子CRISPR関連RNA(scaRNA)及びtracrRNAに依存している。本来、scaRNAは、Cas9を部分的な相補性を有するゲノムDNA標的に誘導し、転写抑制をもたらすと思われる。scaRNAを、他の遺伝子を抑制するようにリプログラミングすることができ、外因性標的に対する操作による相補性の拡張により、DNA切断を誘導することもできる(非特許文献11)。scaRNAがcrRNAと同様にCRISPR-Casシステム内にコードされていることを考慮すると、CRISPRヌクレアーゼをその意図された標的に導くことができるRNAの事前に知られた唯一の供給源は、CRISPR-Casシステム内にしか由来しないこととなる。
【0005】
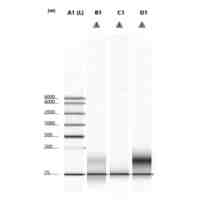
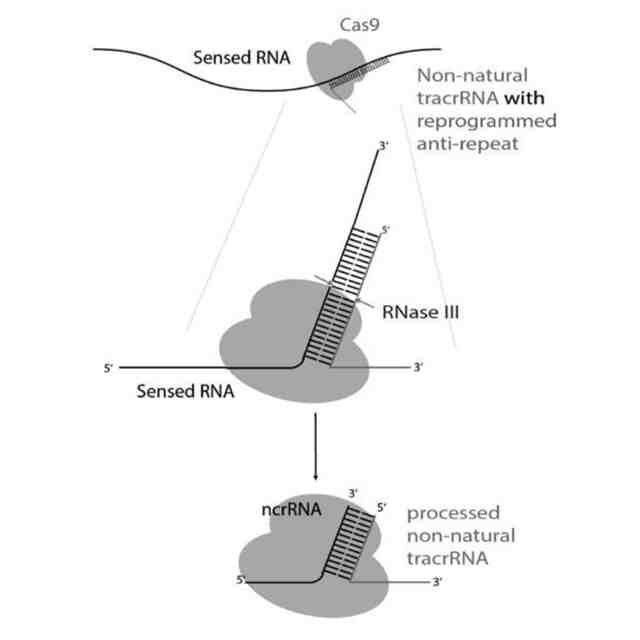
発明者らの最近の出版物(非特許文献12)において、カンピロバクター・ジェジュニ(Campylobacter jejuni)のCas9ヌクレアーゼ(CjeCas9)と共免疫沈降されたRNAがシーケンシングされ、それにより、crRNAのガイド部分に相補性を示すRNAのサブセットがこの細菌のCRISPR遺伝子座内にコードされることが明らかになった。本発明者らは、C.ジェジュニの別個の菌株において同様の共免疫沈降実験を行った。得られた未発表のデータセットには、crRNAとの相補性を示さず、その代わりにtracrRNAのアンチリピート部分との相補性を共有するRNAのセットが含まれていた(図1)。これらのRNAの組成及びサイズは、crRNAの組成及びサイズと似ていた。これらの洞察から、成熟crRNA(Cas9をそれらの標的に導く)が、CRISPRアレイによってコードされたもの以外に、メッセンジャーRNA及び他のRNA(例えば、リボソームRNA、トランスファーRNA、低分子RNA、アンチセンスRNA、核小体低分子RNA、マイクロRNA、piwiRNA、長鎖ノンコーディングRNA、スプライシングされたイントロン、環状RNA)に由来する可能性があることが示唆された。本発明の文脈においては、これらのRNAを、非正規crRNA(ncrRNA)として指定及び/又は呼称する。
【0006】
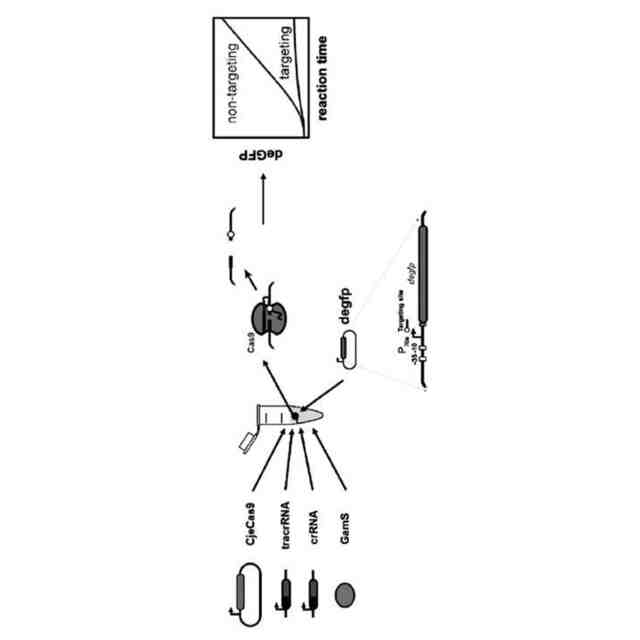
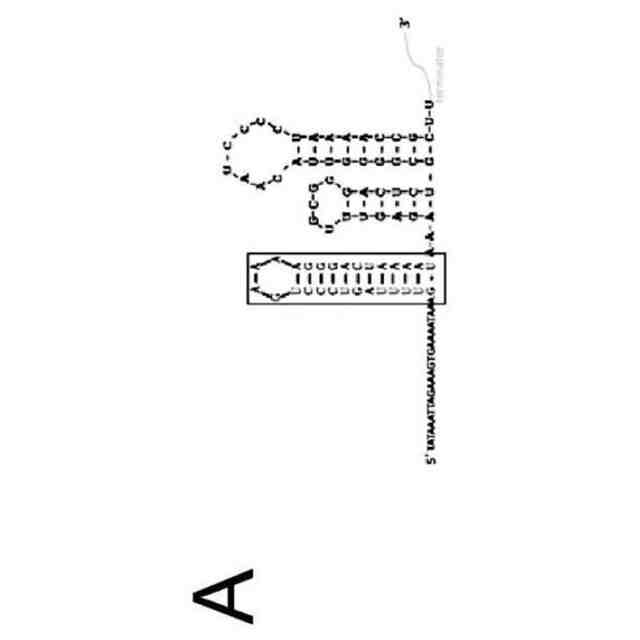
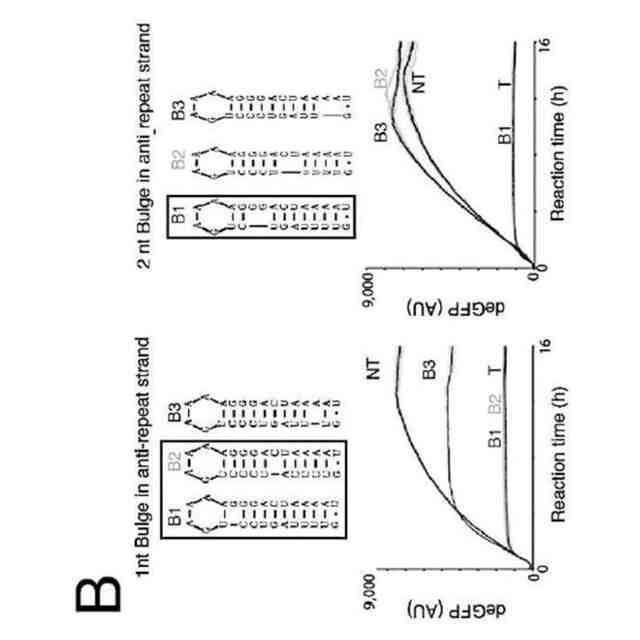
CRISPRリピートとtracrRNAアンチリピートとの間の塩基対合により、tracrRNA依存性CRISPRヌクレアーゼにより結合されるRNA二重鎖が形成される。二重鎖の正確な二次構造は様々であり、二重鎖が完全な相補性を示す場合もあれば、特徴的なバルジを含む場合もある。正確な二次構造にかかわらず、この二次構造をストレプトコッカス・ピオゲネス(Streptococcus pyogenes)由来のCas9のためのsgRNAの部分として維持して、sgRNAのこの領域の配列を改変することができた(非特許文献13)。したがって、本発明者らは、tracrRNAのアンチリピート部分の配列を他のRNAに相補的となるように変化させることにより、各RNAのこの部分が成熟crRNAになり得ると仮定した(図2)。
【0007】
tracrRNAのアンチリピート領域を変化させて、特に任意のRNAをncrRNAに変換するという着想はまだ調査されていない。アンチリピート領域の下流にある領域は、sgRNAの点で改変されており、sgRNAの機能を破壊することなく幾つかの変化に対応することが示されている(非特許文献13)。さらに、Scott, T et al.(非特許文献14)により、アンチリピート領域の外側にある領域を変化させて、Cas9のRNPが示す全体的な編集活性を改善することができることが示されたが、そのメカニズムは不明であった。しかしながら、これらの領域はCas9による認識に関与しており、crRNA又は任意の他のRNAとの対合には関与していない。
【0008】
特許文献1は、tracrRNA配列の少なくとも1つのウラシルヌクレオチドがウラシル以外のヌクレオチドで置き換えられた、遺伝子改変されたトランス活性化型crRNA(tracrRNA)配列を有する核酸配列を利用する組成物及び方法、及び/又はガイドRNA(gRNA)配列を有し、上記gRNA配列の1つ以上のシトシンヌクレオチド及び/又は1つ以上のウラシルヌクレオチドが修飾ヌクレオチドである核酸配列に関する。RNA誘導型DNAエンドヌクレアーゼ酵素、例えば、Cas9又はCpfl又はクラスIIのCRISPRエンドヌクレアーゼ又はそれらの変異体、例えば、不活性化型Cas9(dCas9)又は突然変異型Cas9ニッカーゼ(D10A)を更に含む組成物が開示されている。
【0009】
特許文献2は、合成2パートアプタマー含有ガイドRNAを含む組成物、及び上記合成2パートアプタマー含有ガイドRNAをCRISPR/Casアクチベーターシステムで使用する方法に関する。真核細胞における標的化転写活性化、標的化転写抑制、標的化エピゲノム改変、標的化ゲノム改変、又は標的化ゲノム遺伝子座視覚化の方法が特許請求の範囲に記載されている。
【0010】
特許文献3は、化学的に改変されたcrRNA及びtracrRNA、5'及び/又は3'にコンジュゲートされた部分を有するcrRNA及びtracrRNA、並びにcrRNAのリピート領域又はtracrRNAのアンチリピート領域に改変を有するcrRNA及びtracrRNAを開示している。CRISPRヌクレアーゼを用いるゲノム編集のためにcrRNA及びtracrRNAを使用する方法、並びに上記方法を実施するキットも提供されている。crRNA及びtracrRNAは、培養されたヒト細胞におけるSpyCas9ベースのゲノム編集の有効性を保持しつつ化学的に改変された。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
合同酒精株式会社
麦汁の製造方法
1か月前
池田食研株式会社
RNAの合成方法
2か月前
東洋紡株式会社
細菌からの核酸抽出法
2か月前
杏林製薬株式会社
核酸検出用PCR溶液
今日
熊本県
低褐変レタスとその作製方法
2か月前
東洋紡株式会社
ウイルスからの核酸抽出法
2か月前
学校法人近畿大学
培養肉の製造方法
16日前
JNC株式会社
アデノ随伴ウイルスの精製方法
23日前
テルモ株式会社
液体除去器具
3か月前
国立大学法人山梨大学
受精胚の選別方法及び装置
2か月前
朝日酒造 株式会社
発泡性清酒の製造方法
2か月前
セージ セラピューティクス, インコーポレイテッド
C7、C12、およびC16置換神経刺激性ステロイドおよびそれらの使用方法
1か月前
ヤマト科学株式会社
インキュベータ
16日前
大和ハウス工業株式会社
藻類培養システム
1か月前
日本特殊陶業株式会社
メタン発生抑制装置
1か月前
個人
超音波機能着きウィスキー熟成ボトル用のキャップ
11日前
株式会社アテクト
培養シート
1か月前
東洋紡株式会社
緩衝剤によるヘムタンパク質の安定化方法
11日前
テルモ株式会社
移植用デバイス
3か月前
テルモ株式会社
積層体および移植片の輸送方法
3か月前
株式会社ユーグレナ
観察用遠隔制御装置
3か月前
学校法人慈恵大学
培養皿用の定規
2か月前
住友化学株式会社
細胞チップ
1か月前
日本臓器製薬株式会社
髄核細胞の培養方法
2か月前
住友化学株式会社
細胞用基材
2か月前
住友化学株式会社
細胞用基材
2か月前
三井化学株式会社
作業用装置
2か月前
花王株式会社
肌タイプの分類方法
1か月前
キヤノン株式会社
細胞剥離装置
1か月前
学校法人藤田学園
アンチセンス核酸およびその利用
3日前
国立研究開発法人産業技術総合研究所
細胞観察方法及びその装置
2か月前
ニプロ株式会社
コネクタ及びバッグ
1か月前
花王株式会社
乳酸応答性酵素電極
3か月前
宝酒造株式会社
蒸留酒類、及び蒸留酒類の製造方法
3か月前
学校法人上智学院
生物の管理方法
1か月前
御木本製薬株式会社
ナガミルエキスの製造方法
2か月前
続きを見る
他の特許を見る






 特許ウォッチ
特許ウォッチ