TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025069157
公報種別
公開特許公報(A)
公開日
2025-04-30
出願番号
2025002788,2023115159
出願日
2025-01-08,2019-10-15
発明の名称
TREM2安定化抗体
出願人
ノバルティス アーゲー
代理人
個人
,
個人
,
個人
,
個人
主分類
C07K
16/28 20060101AFI20250422BHJP(有機化学)
要約
【課題】ヒトミエロイド細胞に発現するトリガー受容体2(hTREM2)タンパク質に結合してそれを安定化させる抗体及びこれらの抗体の使用方法を提供する。
【解決手段】hTREM2タンパク質の免疫グロブリンスーパーファミリー(IgSF)ドメインに結合して前記hTREM2タンパク質を安定化させる抗体又はその抗原結合断片を提供する。特定のアミノ酸配列を有する上記抗体又はその抗原結合断片も提供する。これらの抗体又は抗体をコードするベクターは神経炎症性又は神経変性疾患疾患の治療に使用できる。
【選択図】なし
特許請求の範囲
【請求項1】
ヒトミエロイド細胞上トリガー発現2(human triggering expr
ession on myeloid cells 2)(hTREM2)タンパク質の
免疫グロブリンスーパーファミリー(IgSF)ドメインに結合して前記hTREM2タ
ンパク質(例えば配列番号1、2又は3)を安定化させる抗体又はその抗原結合断片。
続きを表示(約 1,500 文字)
【請求項2】
hTREM2のD39、S40、M41、K42、W44、G45、R46、R47、
H67、N68、L69、W70、L71、L72、F74、L75、R77、D87、
T88、L89及びG90からなる群から選択される1つ以上の残基に結合する、請求項
1に記載の抗体又はその抗原結合断片。
【請求項3】
hTREM2のD39、S40、M41、K42、W44、G45、R46及びR47
からなる群から選択される1つ以上の残基、H67、N68、L69、W70、L71、
L72、F74、L75及びR77からなる群から選択される1つ以上の残基、並びにD
87、T88、L89及びG90からなる群から選択される1つ以上の残基に結合する、
請求項1に記載の抗体又はその抗原結合断片。
【請求項4】
hTREM2のD39、S40、M41、K42、W44、G45、R46及びR47
からなる群から選択される5つ以上の残基、H67、N68、L69、W70、L71、
L72、F74、L75及びR77からなる群から選択される4つ以上の残基、並びにD
87、T88、L89及びG90からなる群から選択される3つ以上の残基に結合する、
請求項1に記載の抗体又はその抗原結合断片。
【請求項5】
hTREM2のD39、S40、M41、K42、W44、G45、R46及びR47
からなる群から選択される7つ以上の残基、H67、N68、L69、W70、L71、
L72、F74、L75及びR77からなる群から選択される4つ以上の残基、並びにD
87、T88、L89及びG90からなる群から選択される3つ以上の残基に結合する、
請求項1に記載の抗体又はその抗原結合断片。
【請求項6】
hTREM2のD39、S40、M41、K42、W44、G45、R46及びR47
からなる群から選択される5つ以上の残基、残基H67、N68、L69、W70、L7
1、L72、F74、L75及びR77の全て、並びにD87、T88、L89及びG9
0からなる群から選択される3つ以上の残基に結合する、請求項1に記載の抗体又はその
抗原結合断片。
【請求項7】
hTREM2のS40、M41、W44、G45、W70、L71、L72、F74、
T88及びL89からなる群から選択される1つ以上の残基に結合する、請求項1に記載
の抗体又はその抗原結合断片。
【請求項8】
hTREM2のS40、M41、W44、G45、W70、L71、L72、F74、
T88及びL89からなる群から選択される2つ以上の残基に結合する、請求項1に記載
の抗体又はその抗原結合断片。
【請求項9】
hTREM2のS40、M41、W44、G45、W70、L71、L72、F74、
T88及びL89からなる群から選択される3つ以上の残基に結合する、請求項1に記載
の抗体又はその抗原結合断片。
【請求項10】
hTREM2のS40、M41、W44、G45、W70、L71、L72、F74、
T88及びL89からなる群から選択される4つ以上の残基に結合する、請求項1に記載
の抗体又はその抗原結合断片。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
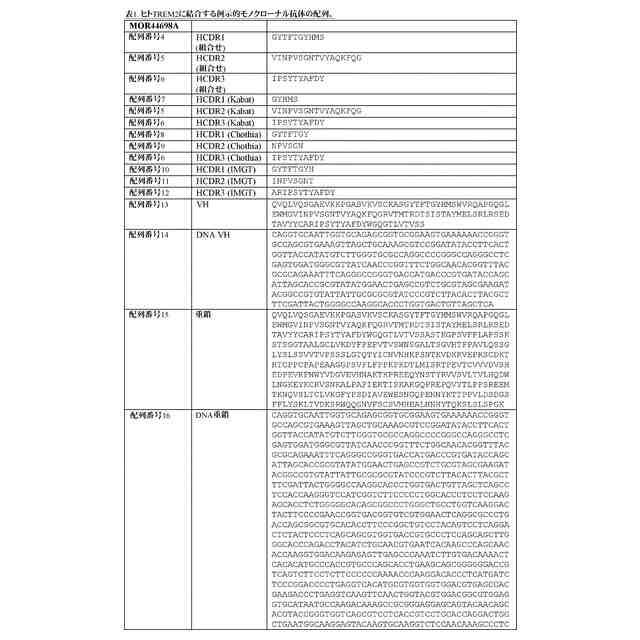
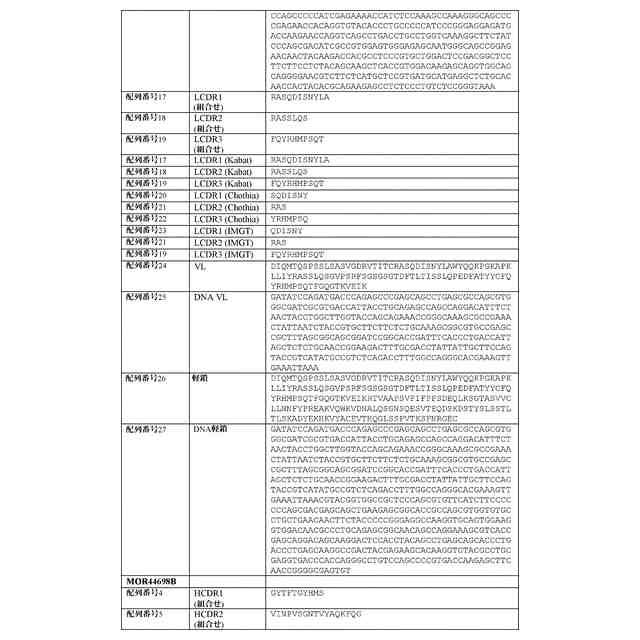
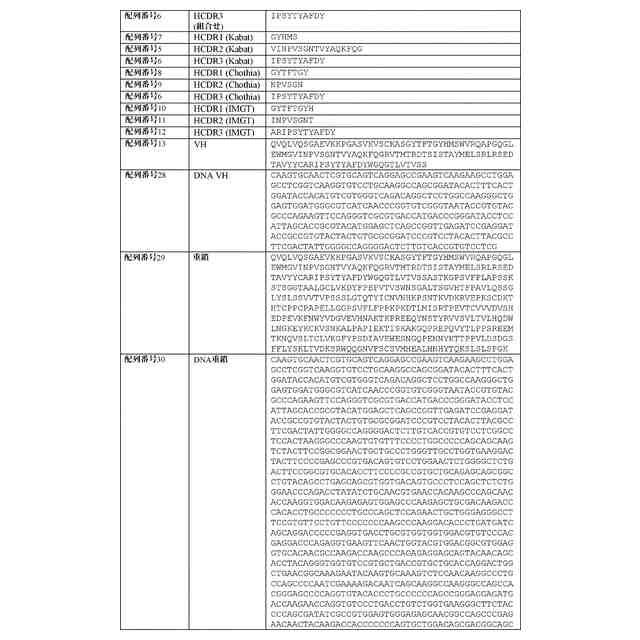
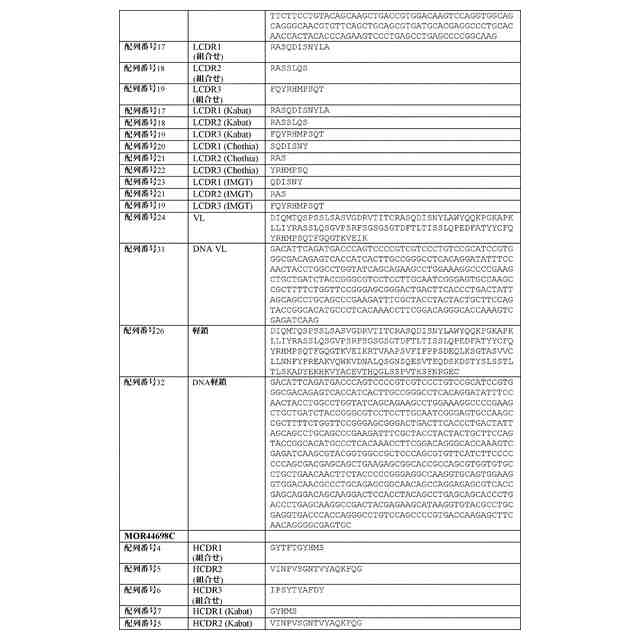
配列表
本出願は、ASCII形式で電子的手段により提出された配列表を含み、その全体が参
照により本明細書に組み込まれる。2019年9月16日に作成された前記ASCIIの
複製は、PAT058251_ST25.txtと命名され、サイズが147,551バ
イトである。
続きを表示(約 4,300 文字)
【0002】
本発明は、ヒトミエロイド細胞に発現するトリガー受容体2(TREM2)タンパク質
に結合してそれを安定化させる抗体及びこれらの抗体の使用方法を提供する。
【背景技術】
【0003】
ミエロイド細胞に発現するトリガー受容体又は「TREM」は、マクロファージ、樹状
細胞、破骨細胞、ミクログリア、マスト細胞、単球、肺上皮細胞、皮膚ランゲルハンス細
胞、クッパー細胞、及び好中球など、様々な種類の骨髄系細胞に発現する一群の膜貫通糖
タンパク質である(Takaki,R.et al.,Immunol.Rev.,20
06,214:118-29)。TREMは、その細胞外ドメインに免疫グロブリン(I
g)型の折り畳みを有し、そのため免疫グロブリンスーパーファミリー(IgSF)に属
する。TREM受容体は短い細胞内ドメインを含むが、シグナル伝達メディエーターのド
ッキングモチーフを欠いており、細胞活性化には、DAP12(12kDaのDNAX活
性化タンパク質)などのアダプタータンパク質が必要である。TREMについては2つの
メンバー、TREM1及びTREM2が報告されており、これらはいずれも免疫及び炎症
反応において重要な役割を果たしている。ヒトTREMをコードする遺伝子は、TREM
1、TREM2、TREM3、TREM4及びTREM5をコードする遺伝子クラスター
、並びにTREM様遺伝子を担持する染色体6p21.1に位置する。
【0004】
TREM2は約40kDaの糖タンパク質であり、これはN-脱グリコシル化後に26
kDaに減少する。TREM2タンパク質全体は、リーディングシグナルペプチド(アミ
ノ酸1~18)、単一V型IgSF細胞外領域(アミノ酸19~132)、ストーク領域
(アミノ酸133~172)、正電荷膜貫通ドメイン(アミノ酸173~197)、及び
サイトゾル尾部(アミノ酸198~230)からなる(Kober et al.,El
ife 5(2016);Kober et al.,J.Mol.Biol.429(
2017)1607-1629)。エキソン2によってコードされる細胞外領域は、3つ
の潜在的N-グリコシル化部位を含む単一のV型IgSFドメインで構成される。推定上
の膜貫通領域は荷電リジン残基を含む。TREM2の細胞質尾部はシグナル伝達モチーフ
を欠いており、シグナル伝達アダプター分子DAP12/TRYROBPを通じてシグナ
ルを送ると考えられている。
【0005】
TREM2はDAP12と物理的に会合し、DAP12は、TREM2及びいくつもの
他の細胞表面受容体のシグナル伝達アダプタータンパク質として働く。DAP12の細胞
質ドメインは免疫受容活性化チロシンモチーフ(ITAM)を含む(Wunderlic
h,J.Biol.Chem.288,33027-33036,2013)。この相互
作用性受容体の活性化後、DAP12には、Srcキナーゼによる保存されたITAMチ
ロシン残基のリン酸化が起こる。続くSykプロテインキナーゼの動員及び活性化により
、マイトジェン活性化プロテインキナーゼ(MAPK)、PI3K、NFκB及びホスホ
リパーゼCγ(PLCγ)の活性化を含め、下流シグナル伝達経路が惹起される。
【0006】
TREM2は、リポ多糖類(LPS)、熱ショックタンパク質60、神経突起破片、細
菌、アポリポタンパク質E及び広範囲のアニオン性及び双性イオン性脂質、例えば、ホス
ファチジン酸(PA)、ホスファチジルグリセロール(PG)、ホスファチジルセリン(
PS)、ホスファチジルイノシトール(PI)、ホスファチジルコリン(PC)、カルジ
オリピン及びスフィンゴミエリンによって活性化し得る。TREM2が活性化すると、ミ
クログリア及びマクロファージの貪食能が増加し、炎症誘発性サイトカインの放出が減少
し、及びTLRシグナル伝達が抑制される。TREM2はCSF-1受容体シグナル伝達
との相乗作用によってミクログリアの生存を維持する。さらに、TREM2はプレキシン
A1と相互作用して細胞接着及び運動性を調節する。TREM2はまた、Aβプラーク又
は神経細胞破片と接触するミクログリア細胞表面領域に濃密に存在する(Yuan et
al.,Neuron 90(2016)724-739)。最近になって、この環境
でTREM2によって感知される一部のリガンド、例えばリン脂質及びミエリン脂質(P
oliani et al.,J.Clin.Invest.125(2015):21
61-2170)並びにApoE(Atagi et al.,J.Biol.Chem
.290(2015):26043-26050;Bailey et al.,J.B
iol.Chem.290(2015):26033-26042)が同定されている。
他のリガンドを挙げるならば、TREM2はミクログリアへのAβの取込みに寄与するた
め、Aβ及びプラーク関連神経細胞破片であり得る(Xiang et al.,EMB
O Mol.Med.8(2016):992-1004)。TREM2はまた、アポト
ーシス細胞(Takahashi et al.,J.Exp.Med.201(200
5),647-657)、ミエリン破片(Poliani et al.,J.Clin
.Invest.125(2015):2161-2170)及び細菌ビーズ(Cen
et al.,Am.J.Respir.188(2013)201-212)の排除に
おいても役割を果たすことが示されている。TREM2シグナル伝達は、取り込まれた被
食物の分解を促進し、脂質代謝、ミエリンの取込み及び細胞内分解に決定的に重要である
。
【0007】
TREM2は、エクトドメインシェディング及び膜内タンパク質分解による逐次的なタ
ンパク質分解プロセシングを受ける(Wunderlich,J.Biol.Chem.
288,33027-33036,2013)。エクトドメインシェディングの間に、A
DAM(ディスインテグリン及びメタロプロテイナーゼドメイン含有タンパク質)又はB
ACE(βサイトAPP切断酵素)ファミリーのメンバーなどのプロテアーゼによってT
REM2のエクトドメインが放出される(Kleinberger,Sci.Trans
l.Med.2014;6(243):243ra86)。
【0008】
エクトドメインが取り除かれた後、残りの膜に保持されている断片がγ-セクレターゼ
媒介性の膜内タンパク質分解によってさらにプロセシングされる。エクトドメインシェデ
ィングによって産生されるTREM2の可溶性断片(sTREM2)が、樹状細胞培養物
の上清中並びに非炎症性神経疾患及び多発性硬化症患者の血漿及びCSF(脳脊髄液)サ
ンプル中に観察されている(Kleinberger,2014)。ヒトCSF中のシェ
ディングされたTREM2エクトドメイン、すなわちsTREM2は、潜在的なアルツハ
イマー病(AD)バイオマーカーとして評価されており、一般に老化過程で増加すること
が示されている(Suarez-Calvet,EMBO Mol.Med.8,466
-476,2016)。ADの経過における詳細な分析により、sTREM2がAD初期
に臨床症状が現れる前に増加し、MCI-ADでピークに達し、AD型認知症では上昇し
たまま、但しMCI-AD段階と比較すると低いレベルに留まることが明らかになった(
Suarez-Calvet,2016)。
【0009】
疾患のピークにおけるTREM2発現の増加は、消散を促す(例えば、腹膜炎、創傷治
癒)(Turnbull,2006;Gawish,2015)。神経炎症のような慢性
炎症条件下では、TREM2は絶えずシェディングされ、ミクログリア及びマクロファー
ジにおいてそのシグナル伝達機能を発揮することができない。従って、細胞表面でのTR
EM2のシェディングを安定化させ及び/又は防止すると、ミクログリア及びマクロファ
ージにおいて機能性のシグナル伝達能を有するTREM2発現が回復することになる。
【0010】
ヒト遺伝学研究では、sTREM2がないことよりむしろ、表面TREM2が失われる
ことが疾患リスクを左右すると指摘されている。例えば、TREM2タンパク質、例えば
配列番号1の47位におけるRからHへのアミノ酸変異は、細胞表面発現の僅かな低下(
Kleinberger 2014)、及びTREM2のリガンド結合能の低下(Wan
g 2015、Atagi 2015、Bailey 2015)を引き起こす。TRE
M2におけるアミノ酸変異T66Mは、細胞表面におけるTREM2発現の欠損をもたら
し(Kleinberger 2014)、ひいては可溶性TREM2が生成されなくな
る。TREM2の切断部位における変異H157Yは、sTREM2の発現を増強して完
全長膜結合型TREM2を減少させ、ADリスクの増加と関連付けられる(Thornt
on 2017、Schlepckow 2017)。従って、これらの遺伝学研究から
、sTREM2の減少及び細胞膜結合型TREM2の増加の両方に細胞表面のTREM2
の安定化が望ましいことが示唆される。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
高砂香料工業株式会社
香料組成物
2か月前
株式会社トクヤマ
グルコンアミド誘導体
17日前
日本化薬株式会社
新規顕色剤及び記録材料
24日前
花王株式会社
ポリアミド系ポリマー
1か月前
日本化薬株式会社
新規顕色剤及び記録材料
24日前
ダイキン工業株式会社
シラン化合物
17日前
日本化薬株式会社
シアノ複素環化合物の製造方法
2か月前
東ソー株式会社
1,2-ジクロロエタンの製造方法
2か月前
個人
メタンガス生成装置およびメタンガス生成方法
2か月前
株式会社トクヤマ
グルコンアミド誘導体の製造方法
17日前
花王株式会社
新規ピリジニウム化合物
23日前
個人
メタンガス生成装置およびメタンガス生成方法
2か月前
株式会社クラレ
メタクリル酸メチルの製造方法
2か月前
株式会社トクヤマ
結晶形Iのリオシグアトの製造方法
1か月前
国立大学法人京都大学
細胞質送達ペプチド
14日前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
1か月前
ダイキン工業株式会社
分離方法
1か月前
日本曹達株式会社
エチルメチルスルホンの製造方法
2か月前
石原産業株式会社
シアノイミダゾール系化合物の製造方法
1か月前
株式会社半導体エネルギー研究所
有機化合物、発光デバイス
1か月前
ダイキン工業株式会社
SF5含有シラン化合物
9日前
小川香料株式会社
化合物及び香料組成物
14日前
小川香料株式会社
化合物及び香料組成物
14日前
石原産業株式会社
シクラニリプロールの製造中間体の製造方法
1か月前
大正製薬株式会社
MMP9阻害作用を有するインドール化合物
2か月前
株式会社アイティー技研
炭化水素の合成方法及び合成装置
2か月前
オリザ油化株式会社
新規化合物及びその用途
1か月前
株式会社トクヤマ
精製第4級アンモニウム化合物水溶液の製造方法
今日
日本化薬株式会社
イソシアヌレート環を有する多官能アミン化合物
3か月前
国立大学法人 東京大学
アシルヒドラゾン誘導体
2か月前
株式会社レゾナック
C2化合物の製造方法
1か月前
キッコーマン株式会社
ナノポアタンパク質
1か月前
ユニマテック株式会社
フェノチアジン誘導体化合物の精製方法
2か月前
国立大学法人九州大学
重水素化化合物の製造方法
1か月前
四国化成工業株式会社
トリアジン化合物、その合成方法およびその利用
1か月前
三洋化成工業株式会社
生体由来材料中のコロイド成分濃度を高める方法
2か月前
続きを見る
他の特許を見る



 特許ウォッチ
特許ウォッチ