TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
公開番号
2025062594
公報種別
公開特許公報(A)
公開日
2025-04-14
出願番号
2024196641,2022052186
出願日
2024-11-11,2014-06-05
発明の名称
RNAガイド遺伝子編集及び遺伝子調節
出願人
デューク ユニバーシティ
代理人
個人
,
個人
,
個人
,
個人
,
個人
,
個人
主分類
C07K
19/00 20060101AFI20250403BHJP(有機化学)
要約
【課題】本明細書では集合等間隔配置短パリンドロームリピート(CRISPR)/CRISPR随伴(Cas)9依拠システムに関連する組成物、及び前記CRISPR/Cas9依拠システム関連組成物を遺伝子発現及びゲノム操作のために使用する方法が開示される。本明細書ではまた、筋肉(例えば骨格筋及び心筋)での遺遺伝発現及び遺伝子操作のための組成物及び前記組成物の使用方法が開示される。
【解決手段】2つの異種ポリペプチドドメインを含む融合タンパク質であって、第一のポリペプチドドメインが、集合等間隔配置短パリンドローム配列リピート随伴(Cas)タンパク質を含み、第二のポリペプチドドメインが、転写活性化活性、転写抑制活性、転写解除因子活性、ヒストン改変活性、ヌクレアーゼ活性、核酸結合活性、メチラーゼ活性、デメチラーゼ活性から成る群から選択される活性を有する、前記融合タンパク質。
【選択図】図1
特許請求の範囲
【請求項1】
2つの異種ポリペプチドドメインを含む融合タンパク質であって、第一のポリペプチドドメインが、集合等間隔配置短パリンドローム配列リピート随伴(Cas)タンパク質を含み、第二のポリペプチドドメインが、転写活性化活性、転写抑制活性、転写解除因子活性、ヒストン改変活性、ヌクレアーゼ活性、核酸結合活性、メチラーゼ活性、デメチラーゼ活性から成る群から選択される活性を有する、前記融合タンパク質。
続きを表示(約 660 文字)
【請求項2】
Casタンパク質がCas9を含む、請求項1に記載の融合タンパク質。
【請求項3】
Cas9が、Cas9のヌクレアーゼ活性をノックアウトする少なくとも1つのアミノ酸変異を含む、請求項2に記載の融合タンパク質。
【請求項4】
前記少なくとも1つのアミノ酸変異がD10A及びH840Aの少なくとも1つである、請求項3に記載の融合タンパク質。
【請求項5】
Cas9タンパク質がiCas9(配列番号:1のアミノ酸36-1403)を含む、請求項3又は4に記載の融合タンパク質。
【請求項6】
第二のポリペプチドドメインが転写活性化活性を有する、請求項1から5のいずれか1項に記載の融合タンパク質。
【請求項7】
第二のポリペプチドドメインが、少なくとも1つのVP16転写活性化ドメインリピートを含む、請求項6に記載の融合タンパク質。
【請求項8】
第二のポリペプチドドメインが、VP16テトラマー(“VP64”)又はp65活性化ドメインを含む、請求項7に記載の融合タンパク質。
【請求項9】
第一のポリペプチドドメインを第二のポリペプチドドメインに結びつけるリンカーをさらに含む、請求項1から8のいずれか1項に記載の融合タンパク質。
【請求項10】
融合タンパク質がiCas9-VP64を含む、請求項1から9のいずれか1項に記載の融合タンパク質。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
関連出願の相互引用
本出願は、米国仮特許出願No.61/831,481(2013年6月5日出願)、米国仮特許出願No.61/839,127(2013年6月25日出願)、米国仮特許出願No.61/904,911(2013年11月15日出願)、米国仮特許出願No.61/967,466(2014年3月19日出願)、及び米国仮特許出願No.61/981,575(2014年4月18日出願)(いずれも参照によりその全体が本明細書に含まれる)に対し優先権を主張する。
政府の所有権に関する記載
本発明は、NIHによって提供された連邦政府グラント番号DP2-OD008586及びR01DA036865並びに米国国立科学財団によって提供されたCBET-1151035により政府の支援を得て達成された。米国政府は本発明に対して一定の権利を有する。
続きを表示(約 4,800 文字)
【0002】
技術分野
本開示は、集合等間隔配置短パリンドロームリピート(CRISPR)/CRISPR随伴(Cas)9-依拠システム及びウイルスデリバリーシステムを用いる、遺伝子発現変更、ゲノム操作及び遺伝子のゲノム変更の分野に関する。本開示はまた、筋肉(例えば骨格筋及び心筋)におけるゲノム操作及び遺伝子のゲノム変更の分野に関する。
【背景技術】
【0003】
哺乳動物系では多くの様々な医学的及び科学的利用(組織再生の刺激、薬剤スクリーニング、遺伝子欠損の補てん、発現停止腫瘍サプレッサーの活性化、幹細胞分化の制御、遺伝子スクリーニングの実施及び合成遺伝子回路の創出を含む)のために、合成転写因子を操作して遺伝子発現を制御してきた。これらの転写因子は内因性遺伝子のプロモーター又はエンハンサーを標的とするか、又はトランスジーン調節のために哺乳動物ゲノムに対して独立した配列を認識するように意図的に設計され得る。ユーザー限定配列を標的とする新規な転写因子を操作するもっとも一般的な方策は、ジンクフィンガータンパク質及び転写アクチベーター様エフェクター(TALF)のDNA結合ドメインに基づく。これらのアプローチは共に、これらのドメインのタンパク質-DNA相互作用の原理を利用して固有のDNA結合特異性を有する新規なタンパク質を操作することを必要とする。これらの方法は多くの利用について広範囲の成功を収めてきたが、タンパク質-DNA相互作用を操作するために必要なタンパク質操作は労力を要し、かつ特殊な専門的技術が要求される。
【0004】
さらに、これら新規なタンパク質は常に有効であるとは限らない。その理由はいまだ不明であるが、タンパク質とゲノム標的部位との結合における後成的改変及びクロマチン状態の影響が関係しているかもしれない。加えて、これらの新規なタンパク質が他の成分と同様に各細胞にデリバーされることを担保することは極めて困難である。これらの新規なタンパク質及びそれらの多種多様な成分をデリバーする既存の方法は別個のプラスミド又はベクターによる細胞へのデリバリーを含み、前記はコピー数の相違のために大いに変動し得る発現レベルをもたらす。さらに、トランスフェクション後の遺伝子活性化はプラスミドDNAの希釈のために一過性であり、一時的な遺伝子発現は治療効果を生じるには十分ではない。さらにまた、このアプローチは、トランスフェクションが容易でない細胞タイプには馴染まない。したがって。これら新規なタンパク質の別の制限は転写活性化の有効性である。
【0005】
位置特異的ヌクレアーゼを用いて、標的のゲノム遺伝子座に位置特異的二本鎖切断を導入することができる。このDNA切断は天然のDNA修繕機構を刺激し、2つの可能な修繕経路の1つをもたらす。ドナー鋳型が存在しないとき、切断は非相同性末端接合(NHEJ)(DNAの小さな挿入又は欠失をもたらすエラー多発修繕経路)によって修繕される。この方法を用いて、標的遺伝子配列のリーディングフレームを意図的に破壊、欠失又は改変させることができる。しかしながら、ヌクレアーゼとともにドナー鋳型が提供されると、細胞性機構は相同組換えによって切断を修繕するであろう(相同組換えはDNA切断の存在下で数桁強化される)。この方法を用いて標的部位のDNA配列に特異的な変化を導入できる。操作されたヌクレアーゼが、多様なヒト幹細胞及び細胞株での遺伝子編集又はマウス肝での遺伝子編集のために用いられてきた。しかしながら、これら技術の実施のための主要なハードルは、効果的で効率的でかつ首尾よいゲノム改変を促進する態様での特定組織へのin vivoデリバリーである。
【0006】
遺伝的な遺伝子疾患は米国の小児に熾烈な影響を与えている。これらの疾患はこれまでのところ治癒をもたらさず、症候を緩和する試みによって何とか切り抜けることができるだけである。数十年間で、遺伝子治療分野はこれらの疾患に対して治癒の見込みをもたらした。しかしながら、治療用遺伝子の細胞及び患者への安全で効率的なデリバリーに関する技術的ハードルがこのアプローチを制限している。デュシェンヌ型筋ジストロフィー(DMD)はもっとも一般的な単一遺伝子疾患であり、3500人の男性の1人に発生する。DMDはジストロフィン遺伝子の遺伝性又は偶発性変異の結果である。ジストロフィンは、筋肉細胞の保全及び機能の調節に必要なタンパク質複合体の不可欠成分である。DMD患者は典型的には小児期に彼ら自身を物理的に支える能力を失い、脆弱化は十代の間に進行し、二十代で死に至る。これまでのDMDのための実験的な遺伝子治療策は、一時的遺伝子デリバリーベヒクルの反復投与を必要とするか、又は外来遺伝物質のゲノムDNAへの永久的組込みを当てにする。これらの方法には共に重大な安全性への懸念がある。さらにまた、これらの方策は大きくかつ複雑なジストロフィン遺伝子配列のデリバリー不能によって制限されてきた。
【発明の概要】
【課題を解決するための手段】
【0007】
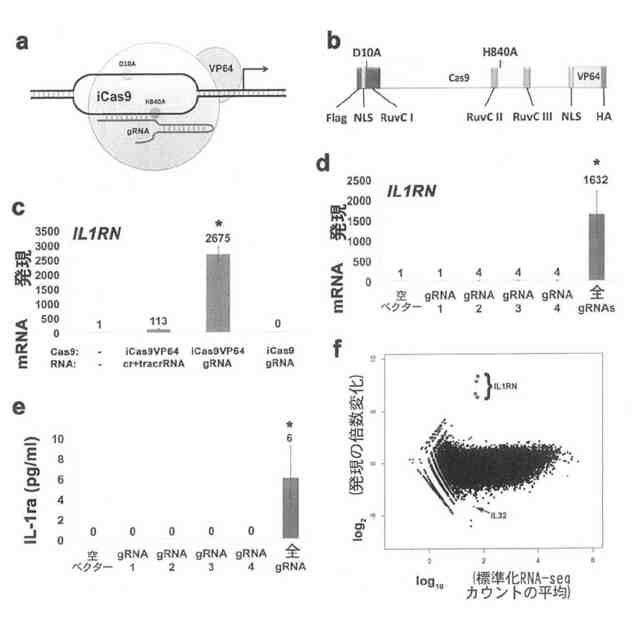
本発明は2つの異種ポリペプチドドメインを含む融合タンパク質を目的とする。第一のポリペプチドドメインは、集合等間隔配置短パリンドロームリピート随伴(Cas)タンパク質を含み、第二のポリペプチドドメインは以下から成る群から選択される活性を有する:転写活性化活性、転写抑制活性、転写解除因子活性、ヒストン改変活性、ヌクレアーゼ活性、核酸結合活性、メチラーゼ活性、デメチラーゼ活性。Casタンパク質はCas9を含むことができる。該Cas9は、Cas9のヌクレアーゼ活性をノックアウトする少なくとも1つのアミノ酸変異を含むことができる。該少なくとも1つのアミノ酸変異は、D10A及びH840Aの少なくとも1つであり得る。該Casタンパク質はiCas9(配列番号:1のアミノ酸36-1403)を含むことができる。第二のポリペプチドドメインは転写活性化活性を有することができる。第二のポリペプチドドメインは、少なくとも1つのVP16転写活性化ドメインリピートを含むことができる。第二のポリペプチドドメインは、VP16テトラマー(“VP64”)又はp65活性化ドメインを含むことができる。融合タンパク質はさらに、第一のポリペプチドドメインを第二のポリペプチドドメインに結びつけるリンカーを含むことができる。融合タンパク質はiCas9-VP64を含むことができる。
【0008】
本発明は、前記融合タンパク質及び少なくとも1つのガイドRNA(gRNA)を含むDNAターゲティングシステムを目的とする。少なくとも1つのgRNAは、標的DNA配列の12-22塩基対の相補性ポリヌクレオチド配列とその後に続くプロトスペーサー隣接モチーフを含むことができる。当該少なくとも1つのgRNAは、遺伝子のプロモーター領域、遺伝子のエンハンサー領域、又は遺伝子の転写領域を標的とすることができる。少なくとも1つのgRNAは遺伝子のイントロンを標的とすることができる。少なくとも1つのgRNAは遺伝子のエクソンを標的とすることができる。少なくとも1つのgRNAは、ASCL1、BRN2、MYT1L、NANOG、VEGFA、TERT、IL1B、IL1R2、IL1RN、HBG1、HBG2及びMYOD1から成る群から選択される遺伝子のプロモーター領域を標的とすることができる。少なくとも1つのgRNAは、配列番号:5-40、65-144、492-515、540-563及び585-625の少なくとも1つを含むことができる。
本発明は、ジストロフィン遺伝子と結合する、Cas9及び少なくとも1つのガイドRNA(gRNA)を含むDNAターゲティングシステムを目的とする。少なくとも1つのgRNAはジストロフィン遺伝子のイントロンを標的とすることができる。少なくとも1つのgRNAはジストロフィン遺伝子のエクソンを標的とすることができる。少なくとも1つのガイドRNAは、配列番号:5-40、65-144、492-515、540-563及び585-625の少なくとも1つを含むことができる。DNAターゲティングシステムは1つから10の異なるgRNAを含むことができる。
本発明は、前記融合タンパク質又は前記DNAターゲティングシステムをコードする単離ポリヌクレオチドを目的とする。
本発明は前記単離ポリヌクレオチドを含むベクターを目的とする。
本発明は、前記単離ポリヌクレオチド又は前記ベクターを含む細胞を目的とする。
【0009】
本発明は細胞で哺乳動物遺伝子の発現を調整する方法を目的とする。前記方法は、細胞を前記融合タンパク質、前記DNAターゲティングシステム、前記単離ポリヌクレオチド、又は前記ベクターと接触させる工程を含む。遺伝子発現は誘導することができる。
本発明は、細胞をトランス分化させるか又は細胞の分化を誘発する方法を目的とする。前記方法は、細胞を前記融合タンパク質、前記DNAターゲティングシステム、前記単離ポリヌクレオチド、又は前記ベクターと接触させる工程を含む。細胞は線維芽細胞又は誘導多能性幹細胞であり得る。線維芽細胞はニューロン細胞又は筋原性細胞にトランス分化させ得る。DNAターゲティングシステムを細胞と接触させることができ、少なくとも1つのgRNAは、ASCL1、BRN2、MYOD1及びMYT1Lから成る群から選択される少なくとも1つの遺伝子のプロモーター領域を標的とする。DNAターゲティングシステムは、ASCL1遺伝子のプロモーター領域を標的とする少なくとも1つのgRNA、及びBRN2遺伝子のプロモーター領域を標的とする少なくとも1つのgRNAを含むことができる。DNAターゲティングシステムは1つから20の異なるgRNAを含むことができる。DNAターゲティングシステムは8から16の異なるgRNAを含むことができる。DNAターゲティングシステムはdCas9-VP64を含むことができる。DNAターゲティングシステムは細胞にウイルス的に又は非ウイルス的にデリバーされ得る。
【0010】
本発明は細胞で変異遺伝子を修正する方法を目的とする。前記方法は、前記DNAターゲティングシステム、前記単離ポリヌクレオチド又は前記ベクターを含む細胞に投与する工程を含む。変異遺伝子の修正は相同性指向修繕(homology-directed repair)を含むことができる。前記方法はさらに細胞にドナーDNAを投与する工程を含むことができる。変異遺伝子は、未成熟終止コドン及び切端遺伝子生成物を生じさせるフレームシフト変異を含むことができる。変異遺伝子の修正は、ヌクレアーゼ媒介非相同性末端接合を含むことができる。変異遺伝子の修正は、未成熟終止コドンの欠失、スプライスアクセプター部位の破壊、1つ以上のエクソンの欠失又はスプライスドナー配列の破壊を含むことができる。1つ以上のエクソンの欠失はリーディングフレームの修正をもたらすことができる。
本発明は、その必要がある変異ジストロフィン遺伝子保有対象動物を治療する方法を目的とする。前記方法は、対象動物に前記DNAターゲティングシステム、前記単離ポリヌクレオチド又は前記ベクターを投与する工程を含む。対象動物はデュシェンヌ型筋ジストロフィーを罹患し得る。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
高砂香料工業株式会社
香料組成物
2か月前
株式会社トクヤマ
グルコンアミド誘導体
2日前
花王株式会社
ポリアミド系ポリマー
22日前
日本化薬株式会社
新規顕色剤及び記録材料
9日前
日本化薬株式会社
新規顕色剤及び記録材料
9日前
日産化学株式会社
ピリジン化合物の製造方法
2か月前
トヨタ自動車株式会社
メタン製造装置
2か月前
日本化薬株式会社
シアノ複素環化合物の製造方法
2か月前
ダイキン工業株式会社
シラン化合物
2日前
ダイキン工業株式会社
シラン化合物
2か月前
株式会社トクヤマ
グルコンアミド誘導体の製造方法
2日前
花王株式会社
新規ピリジニウム化合物
8日前
JNC株式会社
ジアミンおよびこれを用いた重合体
2か月前
個人
メタンガス生成装置およびメタンガス生成方法
2か月前
東ソー株式会社
1,2-ジクロロエタンの製造方法
2か月前
個人
メタンガス生成装置およびメタンガス生成方法
2か月前
株式会社クラレ
メタクリル酸メチルの製造方法
2か月前
株式会社トクヤマ
結晶形Iのリオシグアトの製造方法
1か月前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
22日前
日本曹達株式会社
エチルメチルスルホンの製造方法
2か月前
ダイキン工業株式会社
分離方法
1か月前
石原産業株式会社
シアノイミダゾール系化合物の製造方法
28日前
三井金属鉱業株式会社
金属化合物含有物
2か月前
株式会社半導体エネルギー研究所
有機化合物、発光デバイス
1か月前
大正製薬株式会社
MMP9阻害作用を有するインドール化合物
2か月前
住友化学株式会社
芳香族複素環化合物の製造方法
2か月前
石原産業株式会社
シクラニリプロールの製造中間体の製造方法
1か月前
オリザ油化株式会社
新規化合物及びその用途
21日前
株式会社アイティー技研
炭化水素の合成方法及び合成装置
1か月前
株式会社フラスク
含ホウ素化合物および有機EL素子
2か月前
日本化薬株式会社
イソシアヌレート環を有する多官能アミン化合物
2か月前
株式会社レゾナック
C2化合物の製造方法
1か月前
キッコーマン株式会社
ナノポアタンパク質
1か月前
国立大学法人九州大学
重水素化化合物の製造方法
1か月前
ユニマテック株式会社
フェノチアジン誘導体化合物の精製方法
2か月前
国立大学法人 東京大学
アシルヒドラゾン誘導体
1か月前
続きを見る
他の特許を見る
 特許ウォッチ
特許ウォッチ