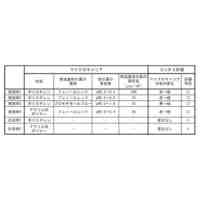
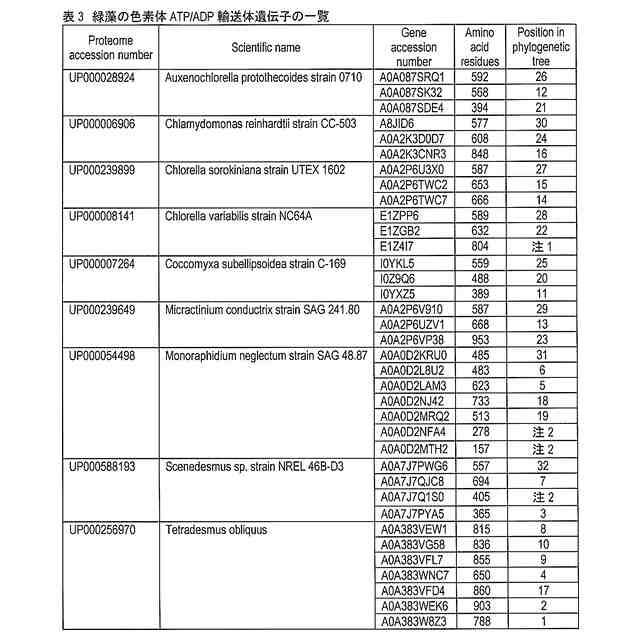
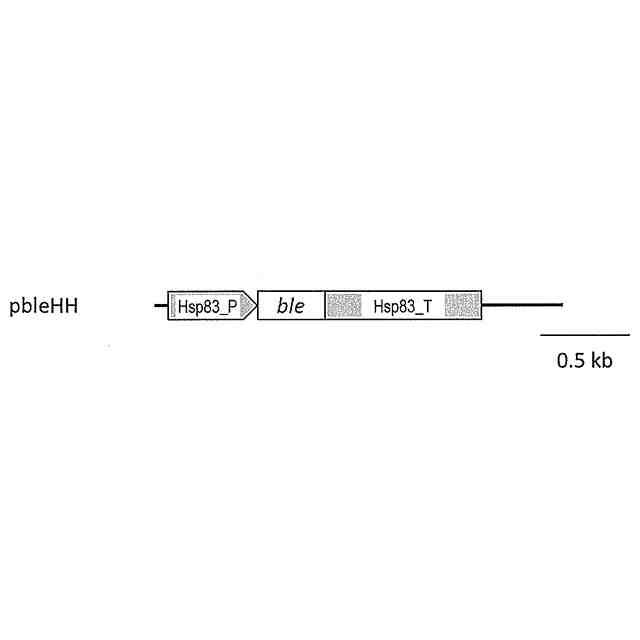
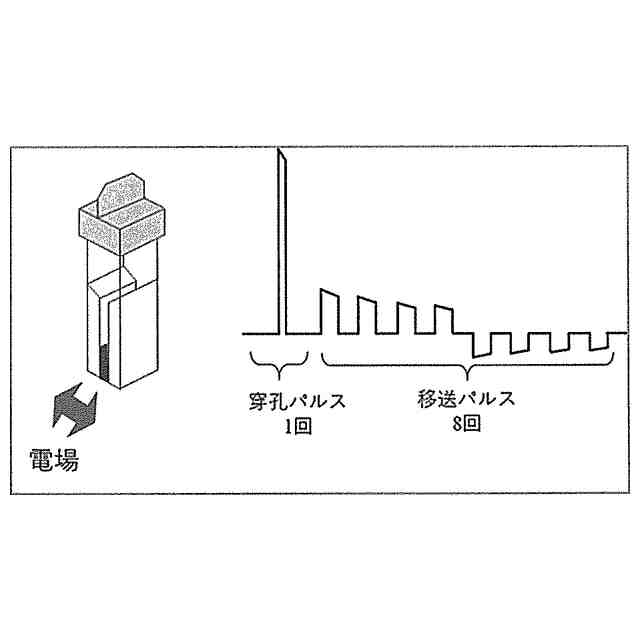
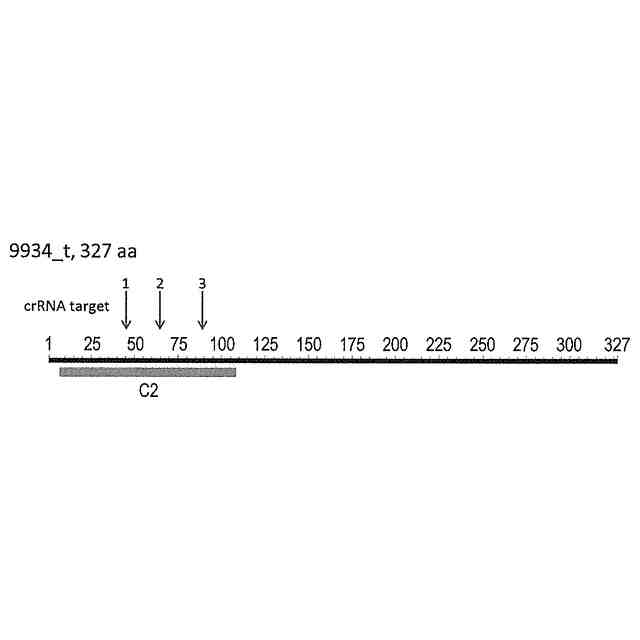
発明の詳細な説明【技術分野】 【0001】 本発明は、油脂含有率及び油脂生産性が増加した緑藻変異体及びその利用に関する。 続きを表示(約 5,500 文字)【背景技術】 【0002】 地球温暖化ガス削減の方策の一つとして、バイオ燃料の開発が挙げられる。その際、食糧と競合しないバイオ燃料開発が必要であるとの認識から、非可食部であるセルロースを原料としたバイオエタノールや、微細藻類の油脂を原料として作られるバイオディーゼル燃料の製造が提案され、それぞれ、第二世代、第三世代のバイオ燃料と呼ばれている。しかし、第二世代バイオ燃料の大規模製造は未だ実現に至らず、技術開発及び実証事業が引き続き進められている。 【0003】 微細藻類は耕作不適地での培養が可能であり、食糧との競合は問題とはならない。それに加え、油糧作物よりも油脂生産性が高いため、バイオディーゼル・バイオジェット燃料の原料として注目を浴びた。2007年以降、複数の米国企業が微細藻燃料の工業的製造に乗り出したが、培養コストや培養液から藻体を回収するためのコストが大きいことが判明し、企業の撤退が相次いでいる。すなわち、第三世代バイオマス燃料においては、現状では、実用化への見通しが立っていない。 【0004】 微細藻バイオ燃料の製造コストが高いことが実用化の最大の障壁であることから、その実用化のためには、培養、回収、油脂抽出、燃料化の各ステップにおいて、コスト削減の努力が必要である。その中でも、培養に関わるコストが全コストの7割程度を占めると言われていることから、培養の効率化、低コスト化が実用化に向けた最重要課題と言える。培養コスト削減の第一の手段は、高い油脂生産性を持つ微細藻類を用いることである。一回の培養で得られる油脂の量が増加すれば、それに反比例して培養コストは減少する。 【0005】 微細藻類とは、酸素発生型光合成を行う単細胞あるいは少数の細胞からなる生物の総称で、シアノバクテリア(原核生物)や、珪藻、黄緑藻、紅藻、緑藻などの真核生物を含む。この多様な微細藻類であるが、バイオディーゼル燃料の原料となる油脂(主成分はトリグリセリド)を蓄積する種類は限られている。国際雑誌論文約200報を調査したところ、高い油脂生産性(単位時間あたりの油脂生産量)を示した微細藻のほとんどは、クロレラ類(Chlorella属、Parachlorella属)とイカダモ類(Desmodesmus属,Scenedesmus 属)に近縁な緑藻であった。 【0006】 Parachlorella(パラクロレラ)属は高い油脂生産能力を持つ(非特許文献1)。パラクロレラNIES-2152株(Parachlorella kessleri)(以下、混同の恐れが無い場合には、「野生株」と記す)を重イオンビーム照射することによって突然変異を誘起し、油脂生産性が向上した変異株を分離する研究が行われ、油脂含有率(細胞の乾燥重量あたりの油脂重量)が向上した変異株であるPK4株が分離された(非特許文献2,3)。この油脂高蓄積変異体であるPK4株が持つ突然変異を同定するために、野生株およびPK4株のゲノム塩基配列解析に加え、野生株のトランスクリプトーム解析が行われた(非特許文献4)。そして野生株とPK4株のゲノム塩基配列を比較した結果、遺伝子番号8741, 9067, 9934で示される3つの遺伝子に、それぞれ、アミノ酸置換、欠失、およびナンセンス変異が存在することが確認された(非特許文献4)。このことから、これら3つの遺伝子変異のいずれかによって、PK4株の油脂含有率の向上がもたらされた可能性が示された。しかしながら、パラクロレラには、遺伝子組換えを含む遺伝子操作法の開発は全く行われてこなかったため、これら3つの遺伝子のいずれが、油脂含有率向上の原因遺伝子であるかを同定することが実質不可能であった。 【先行技術文献】 【非特許文献】 【0007】 Li, X., Pribyl, P., Bisova, K., Kawano, S., Cepak, V., Zachleder, V., ... & Vitova, M. (2013). The microalga Parachlorella kessleri--A novel highly efficient lipid producer. Biotechnology and bioengineering, 110(1), 97-107. Ota, S., Matsuda, T., Takeshita, T., Yamazaki, T., Kazama, Y., Abe, T., & Kawano, S. (2013). Phenotypic spectrum of Parachlorella kessleri (Chlorophyta) mutants produced by heavy-ion irradiation. Bioresource technology, 149, 432-438. Takeshita, T., Ivanov, I. N., Oshima, K., Ishii, K., Kawamoto, H., Ota, S., ... & Kawano, S. (2018). Comparison of lipid productivity of Parachlorella kessleri heavy-ion beam irradiation mutant PK4 in laboratory and 150-L mass bioreactor, identification and characterization of its genetic variation. Algal research, 35, 416-426. Ota, S., Oshima, K., Yamazaki, T., Kim, S., Yu, Z., Yoshihara, M., ... & Kawano, S. (2016). Highly efficient lipid production in the green alga Parachlorella kessleri: draft genome and transcriptome endorsed by whole-cell 3D ultrastructure. Biotechnology for biofuels, 9(1), 1-10. Sorokin, C., & Krauss, R. W. (1959). Maximum growth rates of Chlorella in steady-state and in synchronized cultures. Proceedings of the National Academy of Sciences of the United States of America, 45(12), 1740. Neupert, J., Gallaher, S. D., Lu, Y., Strenkert, D., Segal, N. A., Barahimipour, R., ... & Bock, R. (2020). An epigenetic gene silencing pathway selectively acting on transgenic DNA in the green alga Chlamydomonas. Nature communications, 11(1), 1-17. Tatara, H., Sato, Y., Kasai, Y., Hagiwara, D., Makino, A., Imamura, S., ... & Harayama, S. (2020). A method for the preparation of electrocompetent cells to transform unicellular green algae, Coccomyxa (Trebouxiophyceae, Chlorophyta) strains Obi and KJ. Algal Research, 48, 101904. Reinhold, T., Alawady, A., Grimm, B., Beran, K. C., Jahns, P., Conrath, U., ... & Neuhaus, H. E. (2007). Limitation of nocturnal import of ATP into Arabidopsis chloroplasts leads to photooxidative damage. The Plant Journal, 50(2), 293-304. Geigenberger, P., Stamme, C., Tjaden, J., Schulz, A., Quick, P. W., Betsche, T., ... & Neuhaus, H. E. (2001). Tuber physiology and properties of starch from tubers of transgenic potato plants with altered plastidic adenylate transporter activity. Plant Physiology, 125(4), 1667-1678. Linke, C., Conrath, U., Jeblick, W., Betsche, T., Mahn, A., During, K., & Neuhaus, H. E. (2002). Inhibition of the plastidic ATP/ADP transporter protein primes potato tubers for augmented elicitation of defense responses and enhances their resistance against Erwinia carotovora. Plant Physiology, 129(4), 1607-1615. Li, J., Han, D., Wang, D., Ning, K., Jia, J., Wei, L., ... & Xu, J. (2014). Choreography of transcriptomes and lipidomes of Nannochloropsis reveals the mechanisms of oil synthesis in microalgae. The Plant Cell, 26(4), 1645-1665. 【発明の概要】 【発明が解決しようとする課題】 【0008】 真核微細藻類が作る油脂の商業利用が進まない大きな理由として、その製造コストが高いことが挙げられる。この油脂製造コストを削減する手段として、もともと高い油脂生産性を有する微細藻を遺伝的に改良し、より高い油脂生産性を持つ株を作出することが考えられる。それを達成する具体的方策の一つとして、上述のPK4株の、油脂含有率向上という形質をもたらした遺伝子変異を同定すること、そして、それと同等の変異をさまざまな緑藻に導入することにより、それぞれの緑藻の油脂含有率を向上させることが考えられる。また、そのような緑藻変異体を培養することにより、バイオ燃料等に供する油脂製造コストを削減することが可能となる。 【0009】 そこで、本発明は、(1)PK4株が有する油脂含有率を向上させる遺伝子変異を同定し、(2)その遺伝子変異を他の緑藻に導入することにより油脂含有率が向上した緑藻を提供することを目的とする。 【課題を解決するための手段】 【0010】 PK4株が有する3つの変異のいずれが油脂含有率増加という形質を生み出したかを確かめるための複数の方法が存在する。たとえば、PK4株が有する変異遺伝子(これをaで示す)に野生型対立遺伝子(これをAで示す)を導入することにより、この操作で作られた部分二倍体(A/a)の形質が野生株の形質を示した場合に、A遺伝子の劣性変異によって油脂含有率向上というPK4株の形質が獲得されたと結論できる。このような実験を行うためには、試験管内で調製した人工DNAをゲノムに組込ませる技術、すなわち、形質転換体を分離する技術が必要となる。また別の方法としては、野生株の持つ野生型対立遺伝子Aを破壊することによって、破壊株がPK4株の持つ油脂含有率向上という形質を示すようになれば、A遺伝子の変異によって油脂含有率向上というPK4株の形質が獲得されたと結論できる。このような実験を行うためには、ゲノムの特定の遺伝子に変異を導入する技術、例えばゲノム編集技術が必要となる。 (【0011】以降は省略されています) この特許をJ-PlatPat(特許庁公式サイト)で参照する



 特許ウォッチ
特許ウォッチ