TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025036429
公報種別
公開特許公報(A)
公開日
2025-03-14
出願番号
2024207359,2022145694
出願日
2024-11-28,2016-11-11
発明の名称
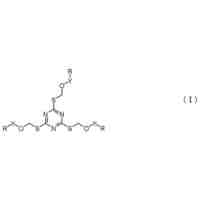
プロテアーゼ活性化受容体2のモジュレーター
出願人
オアシス ファーマシューティカルズ, エルエルシー
,
OASIS PHARMACEUTICALS, LLC
代理人
個人
主分類
C07K
7/08 20060101AFI20250306BHJP(有機化学)
要約
【課題】本明細書において提供されるのは、野生型プロテアーゼ活性化受容体2(PAR2)の変異フラグメントを含むペプチドである。
【解決手段】
疎水性部分を含むペプチドは、細胞膜を透過してPAR2のアンタゴニストとして作用することができる。また本明細書において提供されるのは、当該ペプチドを含む組成物および細胞、ならびに当該ペプチドを使用する方法である。
【選択図】なし
特許請求の範囲
【請求項1】
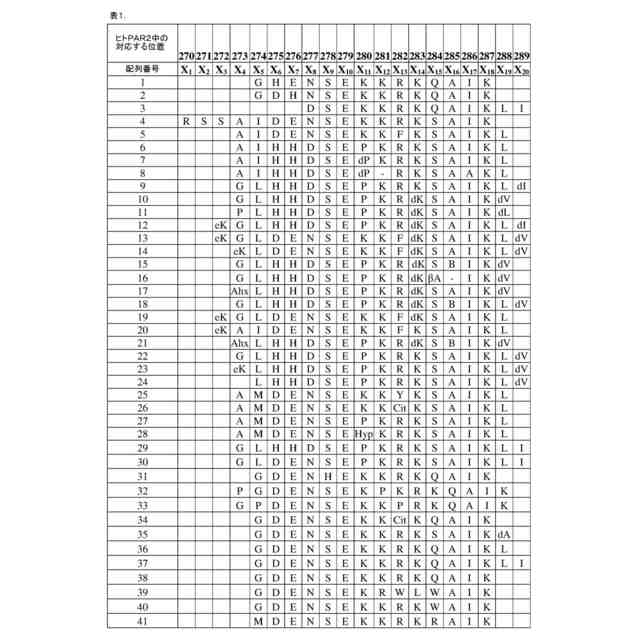
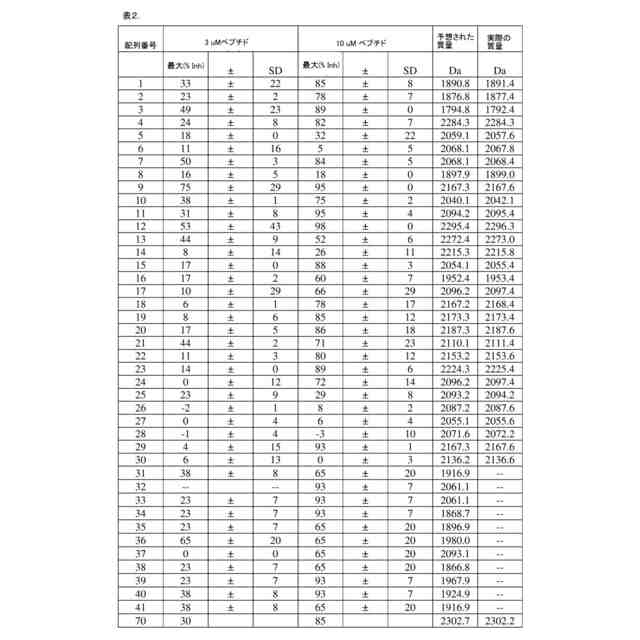
以下の配列:
X
4
X
5
X
6
X
7
X
8
X
9
X
10
X
11
X
12
X
13
X
14
X
15
X
16
X
17
X
18
X
19
X
20
(配列番号42)、
ここで:
X
4
は、不在、A、G、P、またはN末端リンカーであり;
X
5
は、M、G、P、I、L、V、ノルロイシン(J)、メチオニンスルホキシド(M(SO))、またはメチオニンスルホン(M(SO
2
))、またはX
4
が不在である場合は不在であり;
X
6
は、D、E、H、またはX
4
~X
5
が不在である場合は不在であり;
X
7
は、D、E、H、またはX
4
~X
6
が不在である場合は不在であり;
X
8
は、N、D、またはEであり;
X
9
は、任意のアミノ酸であり;
X
10
は、任意のアミノ酸であり;
X
11
は、任意のアミノ酸もしくはそのD-アミノ酸、2-アミノイソ酪酸(B)、ヒドロキシプロリン(Hyp)、P、プロリン相同体、G、または硬化剤/ヘリックス破壊剤部分であり;
X
12
は、K、R、P、または不在であり;
X
13
は、任意のアミノ酸またはシトルリン(Cit)であり;
X
14
は、K、またはX
13
とX
14
との間のペプチド結合を切断不可能にする任意のアミノ酸、または正の電荷を減少させる任意のアミノ酸であり;
続きを表示(約 1,600 文字)
【請求項2】
X
9
が、S、T、H、R、およびKであり;X
10
が、EまたはDであり;およびX
18
が、K、I、またはFである、請求項1に記載のペプチド。
【請求項3】
以下の配列:
X
4
X
5
X
6
X
7
X
8
SEX
11
X
12
X
13
X
14
X
15
X
16
X
17
KX
19
(配列番号43)、
ここで:
X
4
は、不在、A、G、P、またはN末端リンカーであり;
X
5
は、M、G、P、I、L、V、ノルロイシン(J)、メチオニンスルホキシド(M(SO))、またはメチオニンスルホン(M(SO
2
))、またはX
4
が不在である場合は不在であり;
X
6
は、D、E、H、またはX
4
~X
5
が不在である場合は不在であり;
X
7
は、D、E、H、またはX
4
~X
6
が不在である場合は不在であり;
X
8
は、N、D、またはEであり;
X
11
は、任意のアミノ酸もしくはそのD-アミノ酸、2-アミノイソ酪酸(B)、ヒドロキシプロリン(Hyp)、P、プロリン相同体、G、または硬化剤/ヘリックス破壊剤部分であり;
X
12
は、K、R、P、または不在であり;
X
13
は、任意のアミノ酸またはシトルリン(Cit)であり;
X
14
は、KまたはX
13
とX
14
との間のペプチド結合を切断不可能にする任意のアミノ酸、または正の電荷を減少させる任意のアミノ酸であり;
X
15
は、任意のアミノ酸またはベータ-Aであり;
X
16
は、A、S、T、G、Q、ベータ-A、2-アミノイソ酪酸(B)、または不在であり;
X
17
は、I、A、LまたはVであり;および
X
19
は、疎水性アミノ酸、そのD-アミノ酸、または不在である、
【請求項4】
X
13
が、R、F、W、Y、またはシトルリン(Cit)である、請求項1~3のいずれか一項に記載のペプチド。
【請求項5】
X
13
が、Rである、請求項1~4のいずれか一項に記載のペプチド。
【請求項6】
X
13
が、Fである、請求項1~5のいずれか一項に記載のペプチド。
【請求項7】
X
11
が、ヒドロキシプロリン(Hyp)である、請求項1~6のいずれか一項に記載のペプチド。
【請求項8】
X
11
が、Kである、請求項1~6のいずれか一項に記載のペプチド。
【請求項9】
X
11
が、Pである、請求項1~6のいずれか一項に記載のペプチド。
【請求項10】
X
4
が、eK、アミノヘキサン酸(Ahx)、プロリン、またはグリシンからなる群より選択されるN末端リンカーである、請求項1~9のいずれか一項に記載のペプチド。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
関連出願
本願は、2015年11月13日に出願された米国仮出願U.S.S.N. 62/255,334に対する35 U.S.C.§119(e)下における優先権を主張し、当該仮出願は、本明細書において参照することにより援用される。
続きを表示(約 5,600 文字)
【0002】
政府の支援
本発明は、国立衛生学研究所-国立糖尿病消化器腎臓疾患研究所(National Institute of Diabetes and Digestive and Kidney Diseases)により付与された助成金番号R42DK101240下における政府の支援によりなされた。政府は、本発明において一定の権利を有する。
【背景技術】
【0003】
背景
多様なホルモン、神経伝達物質および生物学的に活性な物質は、細胞膜に位置する特異的な受容体を介して、生体の機能を制御、調節または調整する。酵母および哺乳動物を含む真核生物において、これらの受容体のうちの多くは、受容体がカップリングしているグアニンヌクレオチド結合タンパク質(Gタンパク質)を活性化させることにより、細胞内シグナルの伝達を媒介する。かかる受容体は、一般に、Gタンパク質共役受容体(GPCR)と称され、Gタンパク質連動型(G protein-linked)受容体(GPLR)または7回膜貫通型ドメイン受容体(GPCR)としても知られている。GPCRに対する、特異的なシグナル伝達分子、すなわちリガンドの結合は、受容体の立体構造変化を引き起こし得、Gタンパク質に結合してこれを活性化することができる形態をもたらし、それにより、最終的に生物学的応答をもたらす細胞内イベントのカスケードの引き金を引く。典型的には、GPCRは、Gタンパク質と相互作用して、サイクリックAMP、イノシトールリン酸、ジアシルグリセロールおよびカルシウムイオンなどの細胞内セカンドメッセンジャーの合成を調節する。
【0004】
既知の特徴づけられていないGPCRは、それらが多くの疾患に関連するため、薬物の作用および開発の主要な標的となっている(Jacoby et al., Chem. Med. Chem. 2006, 1:760-782)。GPCRは、通常、3つの細胞内(IL-1/i1~IL-3/i3)ループおよび3つの細胞外ループ(EL-1/e1~EL-3/e3)により連結される7つの膜貫通ヘリックスドメイン(TM1~TM7)の共通の構造モチーフを共有する。7つの膜貫通ヘリックスは、細胞膜中にバレル様の空洞を形成し、リガンドとの細胞外相互作用により惹起されるこの構造における立体構造変化こそが、ドメインを細胞内でのGタンパク質共役のためにさらに活性化する。GPCRは、細胞の代謝、線維化、組織リモデリング、細胞の増殖および運動性、接着、炎症、神経型のシグナル伝達、ならびに凝血を制御するシグナル伝達プロセスにおいて、不可欠の役割を果たす。
【0005】
GPCRは、Gタンパク質とエフェクター(Gタンパク質により調節される細胞内の酵素およびタンパク質、ならびにチャネル)と共に、細胞内セカンドメッセンジャーの状態を細胞外の入力に接続するモジュラーシグナル伝達系の成分である(Pierce et al., Nature Rev Mole Cell Bio 2002, 3, 639-650)。GPCRのスーパーファミリーは大きく、ヒトゲノムのシークエンシングにより、それらをコードする850個の遺伝子が明らかとなっている(Hopkins and Groom Nature Reviews Drug Discovery 2002, 1, 727-730)。GPCRは、配列相同性および機能的類似性に基づいて6つのクラスに分けることができる(Foord et al., Pharmacol Rev 2005, 57(2): 279-88);クラスA(または1)(ロドプシン様)、クラスB(または2)(セクレチン受容体ファミリー)、クラスC(または3)(代謝型グルタミン酸/フェロモン)、クラスD(または4)(真菌の接合フェロモン受容体)、クラスE(または5)(サイクリックAMP受容体)、およびクラスF(または6)(Frizzled/Smoothened)。
【0006】
ロドプシン様GPCR(クラスAまたは1)の中には、プロテアーゼ活性化受容体(PAR)があり、これは、7回膜貫通型GPCRのサブファミリーであり、それらの細胞外ドメインの部分の切断を通して活性化され、細胞外プロテアーゼ勾配のセンサーとして作用し、細胞が、線維症、がん、凝血、ならびに無数の他のプロセス(例えば急性および慢性の炎症に関与するものなど)における組織リモデリングの間に、タンパク質分解性微小環境に対して反応することを可能にする。PARファミリーのメンバーは、細胞外プロテアーゼ勾配のセンサーとして作用し、細胞が、組織リモデリングなどの広範な生理学的活動の間に、タンパク質分解性微小環境に反応することを可能にする。今日まで、4つの異なるPAR:PAR1、PAR2、PAR3およびPAR4が同定されている。それらは、ヒトの身体全体にわたり発現される。トリプシン、トロンビン、Xa、VIIa、マトリプターゼ、ヘプシン、トリプターゼおよびMMP-1などのプロテアーゼは、個々のPARメンバーのN末端細胞外ドメインを切断し、それにより、繋留されたリガンドをアンマスクし、これが、受容体の外表面に結合して、細胞内Gタンパク質への膜貫通型シグナル伝達を活性化する。PAR1は、元々、血小板において発見されており、この特殊化されたクラスのGPCRについてのプロトタイプとして役立つ。PAR1は、それが受容体のN末端細胞外ドメイン中に位置する残基R41-S42の間でトロンビンにより切断された場合に、活性化される。PAR3およびPAR4もまた、トロンビンにより活性化されるが、一方、PAR2は、トリプシン/トリプターゼ/Xa/VIIa受容体として最もよく知られている。タンパク質分解による切断は、新たなN末端を露出させ、これが、通常とは異なる細胞内モードにおいて、受容体の本体に結合する。PAR(例えばSFLLRN
PAR1
(配列番号80)、TFLLRN
PAR1
(配列番号71)、PRSFLLRN
PAR1
(配列番号72)、SLIGRL
PAR2
(配列番号73)、AYPGKF
PAR4
(配列番号74)の新たに切断されたN末端 の第1の新しいアミノ酸に対応する合成ペプチドもまた、PARに対する選択的な可溶性の分子間アゴニストとして機能することができる。
【0007】
主要なトロンビン受容体であるPAR1は、内皮バリア機能、血管反応性(vasoreactivity)、内膜過形成、炎症、および恒常性を含む、心血管系の広範な生理学的および病理学的プロセスに影響を及ぼすことが知られている(Ossovskaya et al., Physiol Rev 2004, 84:579-621)。PAR1は、in vitroで内皮細胞の増殖および遊走のメディエーターであり、発達中のマウスにおける血管新生のために必須である。PAR1欠損マウスは、不完全な血管形成のために、妊娠中期(E9.5)において胎仔の半分の死亡をもたらす。驚くべきことに、PAR1欠損マウスは、血小板機能の表現型の変化は有さず、このことが、PAR4の発見につながった。ヒトにおける場合とは異なり、PAR4は、マウスの血小板においては主要なトロンビン受容体であり、PAR4欠損マウスは、トロンビンに対してシグナル応答しない。トリプシン様プロテアーゼの細胞表面受容体であるPAR2は、炎症性細胞、間葉系細胞(例えば線維芽細胞、筋線維芽細胞、平滑筋細胞)、間質細胞、内皮, 肝細胞、星状細胞、ケラチノサイト、膵細胞、神経細胞、心臓細胞、ならびに肺、腸、および肝胆道を含む上皮において広く発現される。PAR2は、皮膚、関節、肺、脳、胃腸管および肝臓、ならびに血管系の多くの急性および慢性の炎症性疾患において重要な役割を果たし、肝臓、肺、腎臓および他の線維性疾患、アトピー性皮膚炎、慢性および急性の疼痛状態、掻痒、および肺動脈性高血圧症の進行に関連付けられている。PAR3の機能的役割は不明であり、合成のPAR3に繋留されたリガンドTFRGAP(配列番号75)は、検出可能な下流のシグナル伝達を刺激しない。PARはまた、機能的なホモダイマー/オリゴマー、およびヘテロダイマー/オリゴマーを形成することが示されている。PAR1およびPAR3は、PAR4のためのコファクターとして役立つことができ、PAR1は、PAR2をトランス活性化することができる(Kaneider et al., Nat. Immunol. 2007, 8:1303-12)。
【0008】
各々のPARは、Gタンパク質の区別し得るサブセットにカップリングする。例えば、PAR1は、Gα-サブユニットG
q
、G
i
およびG
12/13
とカップリングし、これらは、異なるプロテアーゼにより個別的に活性化される。トロンビンは、3つ全てのヘテロ三量体サブユニットを同時に活性化することができ、一方、MMP-1は、より選択的にG
12/13
シグナル伝達を活性化する。PAR1-G
q
は、Ca
2+
を動態化するlnsP
3
のホスホリパーゼC-β生成、およびタンパク質キナーゼC-α(PKCα)を活性化するジアシルグリセロール(DAG)を刺激する。これらが次いで、ホスホリパーゼA
2
およびホスホリパーゼDを活性化する。G
12/13
は、細胞の形状変化、遊走、およびrho依存的な癌化において主要な役割を果たす。PAR2は、Gq、Giおよびベータ-アレスチンシグナル伝達を刺激し得る。先に、PAR1-PAR2ヘテロダイマーに関してG
12/13
からG
i
へのGタンパク質シグナル伝達のスイッチが起こることが、内皮バリア機能の維持に関与していることが示されている。G
i
は、rac、PI3Kの活性化、およびアデニル酸シクラーゼの阻害およびcAMPの抑制に関与している。PAR1およびPAR2が、どのようにしてERK1/2などのMAPキナーゼカスケードのメンバーを調節するかは、まだよく理解されていない。
【発明の概要】
【0009】
PEPDUCINS(商標)と称される細胞浸透性ペプチドは、膜浸透性の疎水性部分を野生型GPCRから誘導されるペプチドに付着させて、それにより、特異的な受容体-Gタンパク質シグナル伝達経路に対する人工のアゴニストおよび/またはアンタゴニストを生成することにより考案された(Covic L. et al. 2002, PNAS 99: 643-48;米国特許第6,864,229号;同第7,696,168号;同第8,389,480号;これらの各々は、本明細書において参照することにより援用される)。これらの脂質付加されたペプチドまたはポリペプチド(「リポペプチド」)は、膜を迅速にフリッピング(flip)するかまたは越えて、高度に特異的な様式において、すなわち、アロステリックな機構によるそれらのコグネートな受容体についての高い選択性により受容体-Gタンパク質シグナル伝達を妨害する能力を有する。PAR、例えばPAR1、PAR2およびPAR4についてのリポペプチド、コレシストキニンAおよびB(CCKA、CCKB)、ソマトスタチン-2(SSTR2)、メラノコルチン-4(MC4R)、グルカゴン様ペプチド-1受容体(GLP-1R)、およびP2Y
12
ADP受容体が作られており、それらが由来する受容体のアゴニストおよび/またはアンタゴニストとして作用する。これらの組成物は、タンパク質ファミリーPARを含む広範なGPCRの活性を活性化または阻害するために有用である。ヒトPARは、PAR1(Genbankアクセッション番号AF019616)、PAR2(Genbankアクセッション番号XM003671)、PAR3(Genbankアクセッション番号NM004101)、およびPAR4(Genbankアクセッション番号NM003950.1)を含み、これらは、本明細書において参照することにより援用される。
【0010】
PEPDUCINs(商標)は、PARファミリーのメンバーについて顕著かつ実質的なアゴニスト効果を有しない効果的なアンタゴニストとして用いられてきたが、一方で、受容体-Gタンパク質共役の機構、および受容体とGタンパク質との間の選択的な接触に対するその関与をさらに研究するため、ならびに、GPCRが関連付けられる多様な疾患および状態の処置および/または予防のための治療剤を作るために、より効果的なPAR2アンタゴニストについての必要性がなお存在する。候補のpepducinsは、疎水性の第2のドメインを、多かれ少なかれ、i3ループなどのインターフェイス接触のいずれかを担う可能性が高いGPCRセグメントと、TM6およびTM5を含むその隣接領域とを含む第1のドメインに付着させること、ならびに/または、受容体中の類似のループ/TMセグメントを潜在的に置き換えるかまたはこれに挿入して、それにより受容体-Gタンパク質シグナル伝達を修飾することにより構築される。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
高砂香料工業株式会社
香料組成物
3か月前
株式会社トクヤマ
グルコンアミド誘導体
27日前
日本化薬株式会社
新規顕色剤及び記録材料
1か月前
日本化薬株式会社
新規顕色剤及び記録材料
1か月前
花王株式会社
ポリアミド系ポリマー
1か月前
日本化薬株式会社
シアノ複素環化合物の製造方法
2か月前
ダイキン工業株式会社
シラン化合物
27日前
東ソー株式会社
1,2-ジクロロエタンの製造方法
3か月前
株式会社トクヤマ
グルコンアミド誘導体の製造方法
27日前
花王株式会社
新規ピリジニウム化合物
1か月前
個人
メタンガス生成装置およびメタンガス生成方法
2か月前
個人
メタンガス生成装置およびメタンガス生成方法
2か月前
株式会社トクヤマ
結晶形Iのリオシグアトの製造方法
2か月前
株式会社クラレ
メタクリル酸メチルの製造方法
3か月前
国立大学法人京都大学
細胞質送達ペプチド
24日前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
1か月前
ダイキン工業株式会社
分離方法
2か月前
日本曹達株式会社
エチルメチルスルホンの製造方法
3か月前
石原産業株式会社
シアノイミダゾール系化合物の製造方法
1か月前
小川香料株式会社
化合物及び香料組成物
24日前
小川香料株式会社
化合物及び香料組成物
24日前
ダイキン工業株式会社
SF5含有シラン化合物
19日前
株式会社半導体エネルギー研究所
有機化合物、発光デバイス
1か月前
石原産業株式会社
シクラニリプロールの製造中間体の製造方法
1か月前
大正製薬株式会社
MMP9阻害作用を有するインドール化合物
3か月前
株式会社アイティー技研
炭化水素の合成方法及び合成装置
2か月前
オリザ油化株式会社
新規化合物及びその用途
1か月前
株式会社トクヤマ
精製第4級アンモニウム化合物水溶液の製造方法
10日前
キッコーマン株式会社
ナノポアタンパク質
2か月前
国立大学法人 東京大学
アシルヒドラゾン誘導体
2か月前
国立大学法人九州大学
重水素化化合物の製造方法
2か月前
株式会社レゾナック
C2化合物の製造方法
1か月前
ユニマテック株式会社
フェノチアジン誘導体化合物の精製方法
3か月前
マナック株式会社
2-ナフタレン誘導体の製造方法
2か月前
四国化成工業株式会社
トリアジン化合物、その合成方法およびその利用
1か月前
国立医薬品食品衛生研究所長
新規ペプチド
1か月前
続きを見る
他の特許を見る



 特許ウォッチ
特許ウォッチ