TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025015536
公報種別
公開特許公報(A)
公開日
2025-01-30
出願番号
2024177862,2022181157
出願日
2024-10-10,2018-05-23
発明の名称
組換えタンパク質
出願人
サイティバ・バイオプロセス・アールアンドディ・アクチボラグ
代理人
個人
,
個人
,
個人
主分類
C07K
1/22 20060101AFI20250123BHJP(有機化学)
要約
【課題】1つには、特にスタフィロコッカス・アウレウス プロテインAのBドメイン及びCドメインに由来する免疫グロブリン結合性タンパク質について、大腸菌及び他のグラム陰性細菌における異種タンパク質の発現を向上させること。
【解決手段】本出願は、1つには、機能性ポリペプチド、及び前記機能性ポリペプチドのN末端に連結された、前記機能性ポリペプチドのシグナルペプチド切断部位とN末端近位構造ユニットとの間のアミノ酸残基の数が14~24個であるような長さを有するN末端スペーサーを含む組換えタンパク質を提供する。
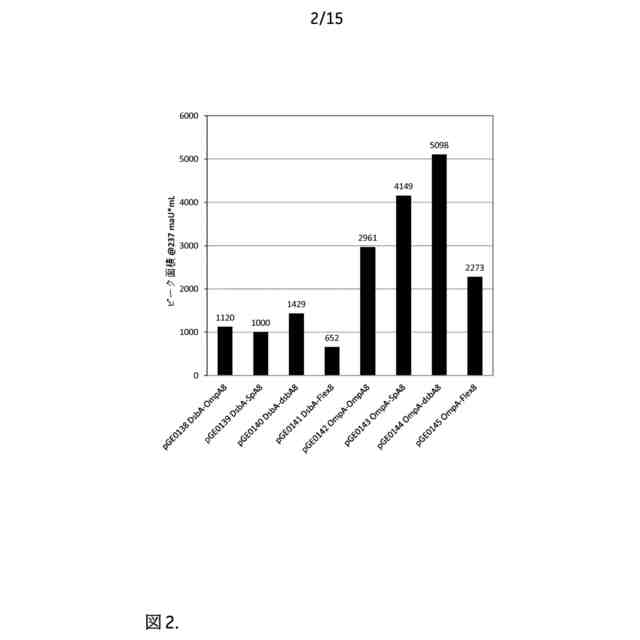
【選択図】図2
特許請求の範囲
【請求項1】
機能性ポリペプチド、及び前記機能性ポリペプチドのN末端に連結された、前記機能性ポリペプチドのシグナルペプチド切断部位とN末端近位構造ユニットとの間のアミノ酸残基の数が14~24個であるような長さを有するN末端スペーサーを含む組換えタンパク質。
続きを表示(約 990 文字)
【請求項2】
免疫グロブリン結合性ポリペプチド、及び前記免疫グロブリン結合性ポリペプチドのN末端に連結された、機能性ポリペプチドのシグナルペプチド切断部位とN末端近位構造ユニットとの間のアミノ酸残基の数が14~24個であるような長さを有するN末端スペーサーを含む組換えタンパク質。
【請求項3】
前記N末端近位構造ユニットがαヘリックスである、請求項1又は2に記載の組換えタンパク質。
【請求項4】
前記機能性又は免疫グロブリン結合性ポリペプチドが少なくとも3つのαヘリックスを含む、請求項3に記載の組換えタンパク質。
【請求項5】
前記N末端スペーサーが、リジン及びアルギニンからなる群から選択される多くとも2個のアミノ酸残基を含む、請求項1から4のいずれか一項に記載の組換えタンパク質。
【請求項6】
前記N末端スペーサーが8~24個のアミノ酸残基からなる、請求項1から5のいずれか一項に記載の組換えタンパク質。
【請求項7】
機能性又は免疫グロブリン結合性ポリペプチド、及び前記機能性又は免疫グロブリン結合性ポリペプチドのN末端に連結された、配列番号16~18、29~30、33~40、43~45、及び47からなる群から選択されるアミノ酸配列と少なくとも80%の配列同一性を有し、又はそれにより規定されるアミノ酸配列を含むN末端スペーサーを含む、組換えタンパク質。
【請求項8】
前記N末端スペーサーが、配列番号16~18、29~30、33~40、43~45、及び47からなる群から選択されるアミノ酸配列と少なくとも80%の配列同一性を有し、又はそれにより規定される、請求項1から7のいずれか一項に記載の組換えタンパク質。
【請求項9】
前記機能性又は免疫グロブリン結合性ポリペプチドが、スタフィロコッカス・アウレウス(Staphylococcus aureus)プロテインAに由来する1つ又は複数のFc結合性ドメインを含む、請求項1から8のいずれか一項に記載の組換えタンパク質。
【請求項10】
前記Fc結合性ドメインがアルカリ安定化Fc結合性ドメインである、請求項9に記載の組換えタンパク質。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明の技術分野
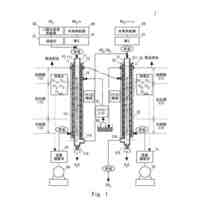
本発明は、組換えタンパク質、特に、大腸菌(Escherichia coli(E. coli))等のグラム陰性細菌において発現した組換えタンパク質に関する。本発明はまた、組換えタンパク質の発現のための核酸、ベクター、及びグラム陰性細菌、加えて、共有結合的に連結された組換えタンパク質リガンドを有する分離マトリックス、及びそのようなマトリックス上で免疫グロブリンを分離する方法に関する。
続きを表示(約 2,900 文字)
【背景技術】
【0002】
本発明の背景
大腸菌(E. coli)における異種タンパク質の発現は、一般的に、実験室規模及び商業的規模での組換えタンパク質について用いられる。分泌を用いる大腸菌での発現は、一般的に、細胞質とペリプラズムを分離する内膜を越えての、産生されたタンパク質の輸送を意味する。ペリプラズムへの分泌により、タンパク質はまた、頻繁に細胞外培地へ漏出する(Mergulhaoら、Biotech Adv 23、177~202頁、2005)。分泌は、細胞質発現と比較して、正しいタンパク質フォールディング、正しいN末端プロセシング、下流処理の単純化、及び封入体への凝集の防止を促進する等の多くの利点を有する。しかしながら、全てのタンパク質がペリプラズムにおける可溶型での発現に成功しているとは言えない。起こり得る問題の一部は、分泌の不足、及びシグナルペプチドのプロセシングがないこと、又は不正確なプロセシングである。不十分な分泌に関する問題を有するタンパク質の特定の例は、スタフィロコッカス・アウレウス(Staphylococcus aureus)のプロテインAの天然又は変異型Fc結合ドメインB又はCに基づいた免疫グロブリン結合体の発現においてである(L Abrahmsenら、EMBO J 4(13B)、3901~3906頁、1985)。そのような免疫グロブリン結合体は一般的に、現代の医薬品の主要なカテゴリーである、モノクローナル抗体のアフィニティークロマトグラフィー分離におけるリガンドとして用いられる。
【0003】
タンパク質発現は、翻訳機構を引きつけるリボソーム結合部位(RBS)、続いて、タンパク質のペリプラズムへの輸送を促進しているシグナルペプチド配列が続く、メッセンジャーリボヌクレオチド酸(ribonucleotide acid)(mRNA)の転写を開始するプロモーター配列に依存する。成熟タンパク質は、シグナルペプチド後にクローニングされる場合が多く、成熟タンパク質は、膜を通過する時、シグナルペプチダーゼによりシグナルペプチドから切り離される。しかしながら、シグナルペプチドの後に構築物をクローニングする時の問題は、制限酵素が、そのDNAを切断するのに特定の配列を必要とする場合が多いことであり、これは、シグナルペプチド配列の後にクローニング跡を残す。
【先行技術文献】
【特許文献】
【0004】
US 8,329,860
US 8,754,196
US 9,040,661
US 9,403,883
JP 2006304633A
US 8,674,073
US 2010/0221844
US 2012/0208234
US 9,051,375
US 2014/0031522
US 2014/0107315
US 2013/0096276
US 2013/0274451
US 2005/0143566
US 2016/0159855
US 2016/0168209
US 2016/0237124
WO 2014/146350
WO 2016/079033
WO 2016/152946
PCT EP2017/061162
PCT EP2017/061164
PCT EP2017/061160
PCT EP2017/061158
PCT EP2017/061159
US 14/961164
US 15/348699
US 15/282367
【非特許文献】
【0005】
Mergulhaoら、Biotech Adv 23、177~202頁、2005
L Abrahmsenら、EMBO J 4(13B)、3901~3906頁、1985
Olsson M.O.及びIsaksson L.A.、Molec. Gen. Genet. 169、251~257頁(1979)
Gel Filtration Principles and Methods、Pharmacia LKB Biotechnology 1991、6~13頁
S Hjerten: Biochim Biophys Acta 79(2)、393~398頁(1964)
「Styrene based polymer supports developed by suspension polymerization"」、R Arshady: Chimica e L'Industria 70(9)、70~75頁(1988)
Paceら、Protein Science 4、2411~2423頁、(1995)
【発明の概要】
【発明が解決しようとする課題】
【0006】
したがって、特にスタフィロコッカス・アウレウス プロテインAのBドメイン及びCドメインに由来する免疫グロブリン結合性タンパク質について、大腸菌及び他のグラム陰性細菌における異種タンパク質の発現を向上させる必要性がある。
【課題を解決するための手段】
【0007】
本発明の概要
本発明の一態様は、大腸菌のようなグラム陰性細菌において容易に発現し、かつ分泌される機能性タンパク質を提供することである。これは、機能性ポリペプチド、及び前記機能性ポリペプチドのN末端に連結された、機能性ポリペプチドのシグナルペプチド切断部位とN末端近位構造ユニットとの間の距離が14~24個のアミノ酸残基であるような長さを有するN末端スペーサーを含む組換えタンパク質で達成される。
【0008】
1つの利点は、発現レベルがN末端スペーサーの導入により向上することである。更なる利点は、シグナルペプチド切断の選択性が向上することである。
【0009】
本発明の第2の態様は、組換えタンパク質をコードする核酸分子を提供することである。これは、5'から3'の方向に以下のエレメントを含む核酸分子で達成され、前記エレメントが作動可能に連結されている:
a)誘導性又は構成的プロモーターDNA配列;
b)シグナルペプチドをコードするDNA配列;
c)N末端スペーサーをコードするDNA配列;及び
d)機能性又は免疫グロブリン結合性ポリペプチドをコードするDNA配列。
【0010】
本発明の第3の態様は、前記の請求項のいずれか一項に記載の組換えタンパク質を発現し、かつグラム陰性細胞の細菌ペリプラズムへ分泌するクローニングベクターを提供することである。これは、上記の核酸分子を含むクローニングベクターで達成される。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
高砂香料工業株式会社
香料組成物
18日前
小野薬品工業株式会社
GCL阻害剤
2か月前
築野グループ株式会社
脂肪酸エステル
3か月前
築野グループ株式会社
脂肪酸エステル
3か月前
日産化学株式会社
ピリジン化合物の製造方法
1か月前
トヨタ自動車株式会社
メタン製造装置
1か月前
ダイキン工業株式会社
シラン化合物
2か月前
日本化薬株式会社
シアノ複素環化合物の製造方法
10日前
ダイキン工業株式会社
シラン化合物
1か月前
個人
メタンガス生成装置およびメタンガス生成方法
10日前
個人
メタンガス生成装置およびメタンガス生成方法
10日前
東ソー株式会社
1,2-ジクロロエタンの製造方法
17日前
田岡化学工業株式会社
多環芳香族化合物の製造方法
2か月前
JNC株式会社
ジアミンおよびこれを用いた重合体
1か月前
学校法人近畿大学
アミロイド線維検出プローブ
3か月前
株式会社クラレ
メタクリル酸メチルの製造方法
19日前
株式会社トクヤマ
ベンジルヒドラジン誘導体の製造方法
2か月前
日本曹達株式会社
エチルメチルスルホンの製造方法
19日前
個人
希土類有機錯体及びその製造方法並び応用
2か月前
三井金属鉱業株式会社
金属化合物含有物
1か月前
株式会社半導体エネルギー研究所
有機金属錯体、発光デバイス
2か月前
大正製薬株式会社
MMP9阻害作用を有するインドール化合物
18日前
住友化学株式会社
芳香族複素環化合物の製造方法
1か月前
日本特殊陶業株式会社
反応装置
3か月前
東ソー株式会社
アデノ随伴ウイルス結合性タンパク質の精製方法
2か月前
日本化薬株式会社
イソシアヌレート環を有する多官能アミン化合物
24日前
株式会社半導体エネルギー研究所
有機化合物、および発光デバイス
2か月前
公立大学法人大阪
尿素化合物の製造方法
1か月前
株式会社フラスク
含ホウ素化合物および有機EL素子
1か月前
グリーンケミカルズ株式会社
4-ヒドロキシ安息香酸組成物
2か月前
東亞合成株式会社
ペプチドフラグメント及びその利用
2か月前
ユニマテック株式会社
フェノチアジン誘導体化合物の精製方法
18日前
国立大学法人富山大学
抗がん作用を有する化合物
3か月前
サンアプロ株式会社
金属錯塩色素
2か月前
ノーベルファーマ株式会社
ヒスチジン亜鉛2水和物の製造方法
2か月前
株式会社Kyulux
有機発光素子
2か月前
続きを見る
他の特許を見る









 特許ウォッチ
特許ウォッチ