TOP
|
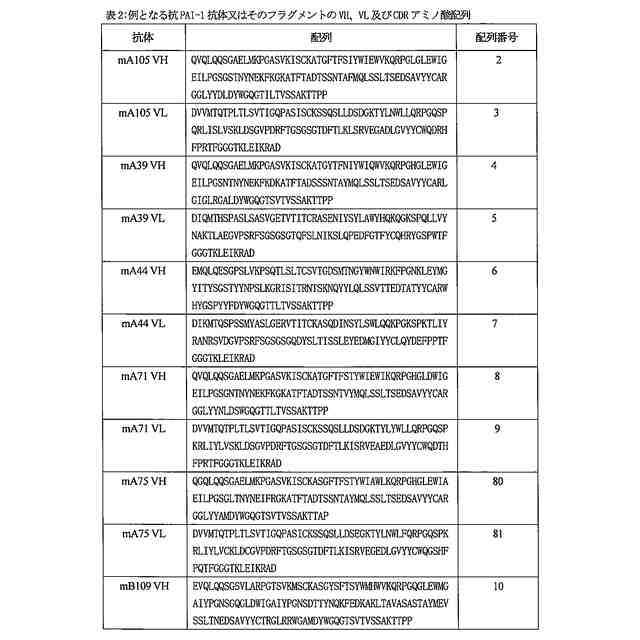
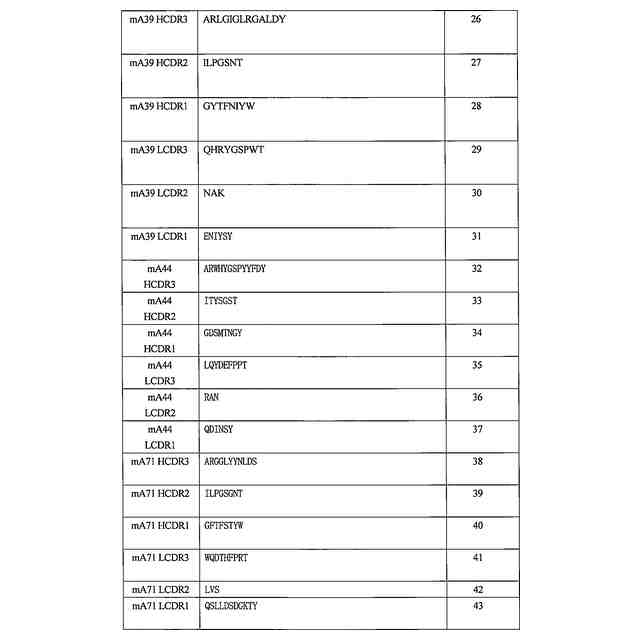
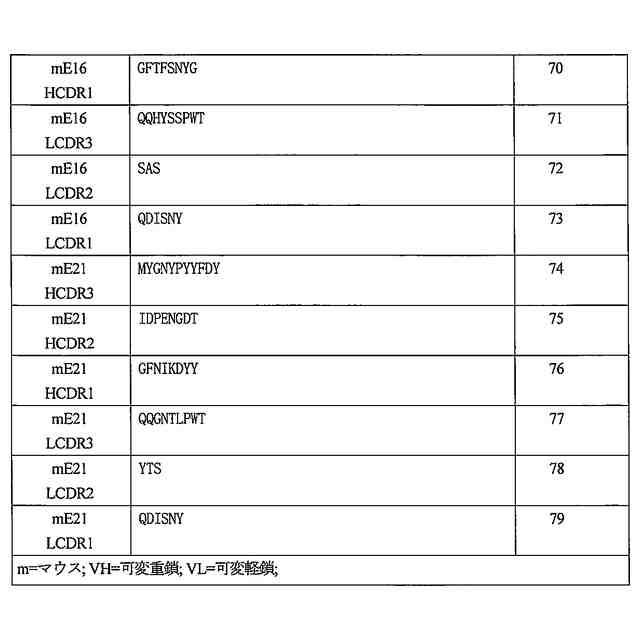
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025011161
公報種別
公開特許公報(A)
公開日
2025-01-23
出願番号
2024172861,2023014335
出願日
2024-10-02,2014-08-13
発明の名称
プラスミノーゲン活性化因子阻害剤-1(PAI-1)に対する抗体及びその使用
出願人
サノフイ
,
SANOFI
代理人
個人
,
個人
主分類
C07K
16/18 20060101AFI20250116BHJP(有機化学)
要約
【課題】プラスミノーゲン活性化因子阻害剤1型(PAI-1)に特異的に結合する抗体、及び該抗体を投与することを含む、プラスミン生成を回復させる方法を提供する。
【解決手段】(a)重鎖フレームワーク領域及び重鎖可変領域、[重鎖可変領域は、特定の配列を含む重鎖CDR1領域、重鎖CDR2領域、重鎖CDR3領域を含む];並びに(b)軽鎖フレームワーク領域及び軽鎖可変領域、[軽鎖可変領域は、特定の配列を含む軽鎖CDR1領域、軽鎖CDR2領域、及び軽鎖CDR3領域を含む]を含む、PAI-1に特異的に結合する単離されたモノクローナル抗体を提供する。
【選択図】なし
特許請求の範囲
【請求項1】
(a) 重鎖フレームワーク領域及び重鎖可変領域、[重鎖可変領域は、配列番号34を含む重鎖CDR1領域、配列番号33を含む重鎖CDR2領域、及び配列番号32を含む重鎖CDR3領域を含む];並びに
(b) 軽鎖フレームワーク領域及び軽鎖可変領域、[軽鎖可変領域は、配列番号37を含む軽鎖CDR1領域、配列番号145を含む軽鎖CDR2領域、及び配列番号35を含む軽鎖CDR3領域を含む]
を含む、PAI-1に特異的に結合する単離されたモノクローナル抗体。
続きを表示(約 3,300 文字)
【請求項2】
(a) 重鎖フレームワーク領域、及び配列番号86を含む重鎖可変領域、並びに
(b) 軽鎖フレームワーク領域、及び配列番号93を含む軽鎖可変領域
を含む、PAI-1に特異的に結合する単離されたモノクローナル抗体。
【請求項3】
(a) 請求項2に記載の抗体の重鎖可変領域と少なくとも95%同一である重鎖可変領域、及び/又は
(b) 請求項2に記載の抗体の軽鎖可変領域と少なくとも95%同一である軽鎖可変領域
を含む、PAI-1に特異的に結合する単離されたモノクローナル抗体。
【請求項4】
請求項1に記載の抗体と本質的に同じエピトープに結合する、単離されたモノクローナル抗体。
【請求項5】
(a) 重鎖フレームワーク領域及び重鎖可変領域、[重鎖可変領域は、配列番号34を含む重鎖CDR1領域、配列番号33を含む重鎖CDR2領域、及び配列番号32を含む重鎖CDR3領域を含む];並びに
(b) 軽鎖フレームワーク領域及び軽鎖可変領域、[軽鎖可変領域は、配列番号37を含む軽鎖CDR1領域、配列番号36を含む軽鎖CDR2領域、及び配列番号35を含む軽鎖CDR3領域を含む]
を含む、PAI-1に特異的に結合する単離されたモノクローナル抗体。
【請求項6】
重鎖可変領域が配列番号6を含み、かつ軽鎖可変領域が配列番号7を含む、請求項5に記載の抗体。
【請求項7】
請求項5に記載の抗体と本質的に同じエピトープに結合する、単離されたモノクローナル抗体。
【請求項8】
(a) 配列番号82を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号91を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント;
(b) 配列番号83を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号92を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント:
(c) 配列番号84を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号93を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント;
(d) 配列番号85を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号91を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント;
(e) 配列番号85を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグ
メント、及び配列番号93を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント;
(f) 配列番号86を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号94を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント;
(g) 配列番号87を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号95を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント;
(h) 配列番号88を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号96を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント;
(i) 配列番号89を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号97を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント;
(j) 配列番号90を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号98を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント;
(1) 配列番号86を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号95を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント;
(m) 配列番号89を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号93を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント;又は
(n) 配列番号89を含む重鎖可変領域を有する重鎖、若しくはその抗原結合フラグメント、及び配列番号95を含む軽鎖可変領域を有する軽鎖、若しくはその抗原結合フラグメント
を含む、ヒトPAI-1に特異的に結合するヒト化モノクローナル抗体。
【請求項9】
(a) 配列番号22を含む重鎖CDR1領域、配列番号21を含む重鎖CDR2領域、及び配列番号20を含む重鎖CDR3領域を含む重鎖可変領域;並びに配列番号25を含む軽鎖CDR1領域、配列番号24を含む軽鎖CDR2領域、及び配列番号23を含む軽鎖CDR3領域を含む軽鎖可変領域、
(b) 配列番号28を含む重鎖CD1領域、配列番号27を含む重鎖CDR2領域、及び配列番号26を含む重鎖CDR3領域を含む重鎖可変領域;並びに配列番号31を含む軽鎖CDR1領域、配列番号30を含む軽鎖CDR2領域、及び配列番号29を含む軽鎖CDR3領域を含む軽鎖可変領域、
(c) 配列番号40を含む重鎖CDR1領域、配列番号39を含む重鎖CDR2領域、及び配列番号38を含む重鎖CDR3領域を含む重鎖可変領域;並びに配列番号43を含む軽鎖CDR1領域、配列番号42を含む軽鎖CDR2領域、及び配列番号41を含む軽鎖CDR3領域を含む軽鎖可変領域、
(d) 配列番号46を含む重鎖CDR1領域、配列番号45を含む重鎖CDR2領域、及び配列番号44を含む重鎖CDR3領域を含む重鎖可変領域;並びに配列番号49を含む軽鎖CD1領域、配列番号48を含む軽鎖CDR2領域、及び配列番号47を含む軽鎖CDR3領域を含む軽鎖可変領域、
(e) 配列番号52を含む重鎖CDR1領域、配列番号51を含む重鎖CDR2領域、及び配列番号50を含む重鎖CDR3領域を含む重鎖可変領域;並びに配列番号55を含む軽鎖CDR1領域、配列番号54を含む軽鎖CDR2領域、及び配列番号53を含む軽鎖CDR3領域を含む軽鎖可変領域、
(i) 配列番号58を含む重鎖CD1領域、配列番号57を含む重鎖CDR2領域、及び配列番号56を含む重鎖CDR3領域を含む重鎖可変領域;並びに配列番号61を含
む軽鎖CD1領域、配列番号60を含む軽鎖CDR2領域、及び配列番号59を含む軽鎖CDR3領域を含む軽鎖可変領域、
(g) 配列番号64を含む重鎖CDR1領域、配列番号63を含む重鎖CDR2領域、及び配列番号62を含む重鎖CDR3領域を含む重鎖可変領域;並びに配列番号67を含む軽鎖CDR1領域、配列番号66を含む軽鎖CDR2領域、及び配列番号65を含む軽鎖CDR3領域を含む軽鎖可変領域、
(h) 配列番号70を含む重鎖CDR1領域、配列番号69を含む重鎖CDR2領域、及び配列番号68を含む重鎖CDR3領域を含む重鎖可変領域;並びに配列番号73を含む軽鎖CDR1領域、配列番号72を含む軽鎖CDR2領域、及び配列番号71を含む軽鎖CDR3領域を含む軽鎖可変領域;又は
(i) 配列番号76を含む重鎖CDR1領域、配列番号75を含む重鎖CDR2領域、及び配列番号74を含む重鎖CDR3領域を含む重鎖可変領域;並びに配列番号79を含む軽鎖CDR1領域、配列番号78を含む軽鎖CDR2領域、及び配列番号77を含む軽鎖CDR3領域を含む軽鎖可変領域
を含む、PAI-1に特異的に結合する単離されたモノクローナル抗体。
【請求項10】
請求項8又は請求項9に記載のヒト化モノクローナル抗体と本質的に同じPAI-1上のエピトープに結合する、PAI-1に特異的に結合する単離されたモノクローナル抗体。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
関連出願の相互参照
本出願は、米国仮出願第61/865,451号(2013年8月13日出願)、及び欧州特許出願第14305757.8号(2014年5月22日出願)(これらはそれら全体として参照により本明細書に加入される)に対する優先権を主張する。
続きを表示(約 4,500 文字)
【背景技術】
【0002】
背景
プラスミノーゲン活性化因子阻害剤-1型(PAI-1)は、プラスミン生成に関与する鍵とな
るセリンプロテアーゼである組織型プラスミノーゲン活性化因子(tPA)及びウロキナーゼ
型プラスミノーゲン活性化因子(uPA)の主要阻害剤である。PAI-1は、血管コンパートメントにおけるプラスミノーゲン活性化を阻害することにより線維素溶解を調節する。線維素溶解は、凝固カスケードの活性化により形成されるフィブリン血栓を分解するための緊密に協調したプロセスである。凝固/線維素溶解バランスの調節不全は、出血又は血栓性疾
患のような異常な止血事象をもたらす。PAI-1はまた、受容体に結合したプラスミノーゲ
ンがウロキナーゼ受容体(uPAR)に結合したウロキナーゼにより主に活性化される細胞周囲コンパートメント(血管内及び組織の)におけるプラスミノーゲン活性化の鍵となる調節因子である。細胞周囲タンパク質分解を阻害することにより、PAI-1は、細胞外マトリッ
クス(ECM)分解、増殖因子活性化及びECMからの放出、マトリックスメタロプロテイナーゼ(MMP)活性化並びに細胞アポトーシスのような多数の細胞昨日を調節する。近年、PAI-1のプロテアーゼ非依存性効果が、(ビトロネクチン、ヘパリン、グリコサミノグリカンのよ
うな)補因子、uPAR-ウロキナーゼ複合体又は細胞受容体(LRP:低密度リポタンパク質受容
体関連タンパク質)又は接着/脱接着、遊走、増殖及び細胞内生物活性のような細胞機能に影響を及ぼすインテグリンとのその相互作用により同定された。これらの細胞機構及び抗線維素溶解効果により、PAI-1の病原性の役割が、腫瘍成長及び転移、線維症、急性心筋
梗塞並びにアテローム性動脈硬化症、肥満及び糖尿病のような代謝障害において確立されてきた。
【0003】
ヒトSERPINE1(PAI-1)遺伝子は、第7染色体に局在化し、8つのイントロン及び9つの
エクソンからなり、そして12,169bのサイズを有する(非特許文献1)。PAI-1は、SERPIN(
セリンプロテアーゼ阻害剤)スーパーファミリーからの約50kDa(379アミノ酸)の単鎖糖タ
ンパク質であり、これは活性コンホメーションで合成されるがビトロネクチン(Vn)が存在しない場合は自発的に潜在性になる。PAI-1の主要補因子であるビトロネクチンは、表面
上に露出された約20個のアミノ酸である反応性中心ループ(RCL)を有する活性コンホメーションを安定化させる。PAI-1の2つの主要な標的(tPA及びuPA)の阻害の機構は自殺阻
害である。PAI-1のRCL領域は、ベイトペプチド結合(R346-M347、P1-P’1とも呼ばれる)を有し、これはこのセリンプロテアーゼについての切断部位を有する。tPA又はuPAとのミカエリス複合体を最初に形成し、次いで触媒三残基はベイトペプチド結合と反応してアシル-酵素複合体を形成し、これがP1-P’1ペプチド結合の切断後に強いコンホメーション変化を誘導する。コンホメーション変化は、プロテアーゼはアシル酵素としてPAI-1と共有結
合したまま、切断されたRCLのβ-ストランドへの挿入を生じる。非生理的環境下で、このアシル-酵素複合体の加水分解は、切断されたPAI-1及び遊離活性プロテアーゼの放出を誘導し得る(非特許文献2)。
【0004】
PAI-1は、非常に変動するレベル(nM範囲)でかつt-PA又はuPA濃度よりも過剰に血中で循環する。PAI-1は構造的柔軟性を示し、そして3つのコンホメーションのうちの1つで見
出され得る:(1) 潜在(latent)コンホメーション、(2)活性コンホメーション、又は(3)基質コンホメーション(図1を参照のこと)。PAI-1は、潜在性移行(latency transition)を1.5~3倍減少させるビトロネクチンとの非共有結合複合体(Kd約1nM)として主に見出され
る。ビトロネクチンに対する潜在的か、切断されたか又は複合体化したPAI-1の親和性は
有意に減少される。マトリックス結合ビトロネクチンもまた、PAI-1とともに細胞周囲の
空間に局在化する。内皮細胞、単球、マクロファージ及び血管平滑筋細胞はこのPAI-1を
合成し、次いでこれが血小板(α顆粒で)により潜在形態で大量に貯蔵され得る。PAI-1は
、溶液中でtPA及びuPAの速く特異的な阻害剤(二次速度定数10
6
~10
7
M
-1
s
-1
を有する)で
あるが、フィブリン又はそれらの細胞受容体のいずれかに結合したプロテアーゼに対して不活性である。トロンビン、プラスミン、活性化プロテインCのような他のプロテアーゼ
もまたPAI-1により阻害されるが効率はより低い。
【0005】
ヒトPAI-1のいくつかの3D構造が、1992年に最初の1つが記載されてから(非特許文献3)、潜在コンホメーションで解析されている。これらの3D構造としては、基質でのPAI-1の変異体形態(非特許文献4)、安定化された活性コンホメーション(非特許文献5)、ビトロネクチン-ソマトメジンBドメインに複合体化したPAI(非特許文献6)又はRCLループからの阻害ペンタペプチドと複合体化したPAI(非特許文献7)が挙げられる。より最近には、潜
在コンホメーションのマウスPAI-1構造が、Dewildeら(非特許文献8)により解明され、そしてRCL位置におけるヒトPAI-1との差異、ゲート領域及びα-ヘリックスAの位置を明らかにした。PAI-1における構造/機能の関係は、この多機能セルピンの様々な活性に関与するドメインの位置を特定するために600より多い変異体タンパク質を使用することにより調べられた(非特許文献9による概説)。
【0006】
PAI-1は、肝細胞、脂肪細胞、メサンギウム細胞、線維芽細胞、筋線維芽細胞、及び上
皮細胞を含むほとんど全ての細胞型により合成され得るので、その発現は、生理的(例えば、血漿PAI-1レベルの概日変動)及び病理的条件(例えば、肥満、代謝症候群、インスリ
ン抵抗性、感染、炎症性疾患、癌)下で大いに変動する。PAI-1は急性期タンパク質と考えられている。PAI-1 mRNA発現の転写調節は、いくつかのサイトカイン及び成長因子(例え
ば、TGFβ、TNFα、EGF、FGF、インスリン、アンギオテンシンII及びIV)、ホルモン(例えば、アルドステロン、グルココルチコイド、PMA、高グルコース)並びにストレス因子(例
えば、低酸素、反応性酸素種、リポ多糖類)により誘導される。
【0007】
さらに、PAI-1遺伝子のプロモーター(位置-675)における多型は発現レベルに影響を及ぼす。4G対立遺伝子はPAI-1レベルを増加させ、そして4G/4G変異体(集団の約25%において発生する)は、5G/5G(25%発生及び4G/5G 50%発生)と比較して血漿PAI-1レベルの約2
5%の増加を誘導する。4G/4G多型は、心筋梗塞(非特許文献10)、特定の型の肺線維症(特発性間質性肺炎)(非特許文献11)に関連付けられており、そして4G/4G遺伝子型ドナー群は、間質性線維症及び尿細管萎縮に起因する移植腎喪失についての独立した危険因子である(非特許文献12)。
【0008】
いくつかの病原性の役割が、動脈血栓症及び静脈血栓症、急性心筋梗塞、及びアテローム性動脈硬化症のような血栓性疾患においてPAI-1に起因すると考えられてきた。インス
リン抵抗性症候群及び肥満のような代謝障害へのその関与はよく認識されている。PAI-1
はいくつかの器官の線維化促進(profibrotic)因子としても知られており、線維性組織
において過剰発現されることが示されている(すなわち、肝臓、肺、腎臓、心臓、腹部癒
着、皮膚:瘢痕又は強皮症)(非特許文献13により概説される)。PAI-1ノックアウト(KO)
マウスは、肝臓(胆管結紮又は生体異物)、腎臓(一側尿管閉塞モデル(UUO))、肺(ブレオマイシン吸入)のような様々なモデルにおいて線維症から保護されるが(非特許文献14;非
特許文献15;非特許文献16)、一方で心臓においてこの欠失は誘導された線維症から保護されるが(非特許文献17)、年齢依存性心選択的線維症になりやすい(非特許文献18)
。siRNAによるPAI-1発現の下方調節(非特許文献19)又は化学化合物による阻害(非特許
文献20;非特許文献21)は、肺線維症を減少させると報告されたが、一方で野生型のPAI-1過剰発現(非特許文献22)又はビトロネクチン結合のみを保持するがtPA阻害剤機能を保持しないPAI-1変異体は肺線維症を増悪させる(非特許文献23)。
【0009】
胆管結紮(BDL)肝線維症は、PAI-1を中和する抗体により減弱されるが(特許文献1)、一方でsiRNAにより下方調節はBDL及び生体異物誘導肝線維症を減弱させる(非特許文献24)。PAI-1 KOマウスは、BDLにおいて胆汁うっ滞性誘導肝損傷及び線維症から(非特許文献25;非特許文献26;非特許文献27)、そしてアンジオテンシンII誘導肝線維症(非特許文献28)から保護された。
【0010】
PAI-1 KOマウスは、UUOモデル(非特許文献29)において、糖尿病性腎症において(非特許文献30)及びアンジオテンシンII誘導腎症において(非特許文献31;概説については
、非特許文献32及び非特許文献33を参照のこと)腎線維症から保護される。対照的に
、PAI-1過剰発現マウスは、より重症の線維症及びUUO後の増加したマクロファージ動員を示す(非特許文献34;非特許文献35)。非阻害性(Non-inhibitory)PAI-1変異体(PAI-1
R)は、ラットにおける実験的糸球体腎炎(thy1)において、尿タンパク質発現及び糸球体
マトリックス蓄積を減少させることにより線維症の発生からマウスを保護することが示された(非特許文献36)。PAI-1をブロックするペプチドは、コラーゲン3、4及びフィブロ
ネクチン蓄積をUUOマウスにおいて阻害する(非特許文献37)。
【先行技術文献】
【特許文献】
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
サノフイ
薬物送達デバイス
20日前
サノフイ
抗CHIKV抗体およびその使用
1か月前
サノフイ
注射デバイスに取り付けるセンサデバイス
1か月前
サノフィ パスツール インコーポレイテッド
ワクチンを設計するためのシステムおよび方法
20日前
サノフィ・バイオテクノロジー
炎症症状で使用するための抗IL-1R3抗体
1か月前
サノフイ
抗原性多量体呼吸器合胞体ウイルスポリペプチド
15日前
サノフイ
B型肝炎ウイルス感染を治療するための併用療法
1か月前
サノフイ
三重特異性結合性タンパク質、方法、およびその使用
15日前
サノフイ
多特異性結合タンパク質において鎖誤対合を分析する方法
7日前
サノフィ・バイオテクノロジー
IL-4Rアンタゴニストを投与することにより喘息を処置又は予防するための方法
1か月前
サノフィ・バイオテクノロジー
IL-4Rアンタゴニストの投与により鼻ポリープを伴う慢性副鼻腔炎を処置する方法
15日前
サノフイ
NKp46およびCD123に結合する多機能性ナチュラルキラー(NK)細胞エンゲージャ
15日前
ニューロクライン バイオサイエンシーズ,インコーポレイテッド
4-(2-クロロ-4-メトキシ-5-メチルフェニル)-N-[(1S)-2-シクロプロピル-1-(3-フルオロ-4-メチルフェニル)エチル]-5-メチル-N-プロパ-2-イニル-1,3-チアゾール-2-アミンの調製のための合成法
1か月前
コニカミノルタ株式会社
フィルム検査装置、フィルム検査方法及びプログラム
1か月前
ケルバー・テクノロジーズ・ゲゼルシャフト・ミト・ベシュレンクテル・ハフツング
ロッド状の物品を取り扱う装置及びこのような装置を初期充填する方法
1か月前
株式会社トクヤマ
グルコンアミド誘導体
16日前
花王株式会社
ポリアミド系ポリマー
1か月前
日本化薬株式会社
新規顕色剤及び記録材料
23日前
日本化薬株式会社
新規顕色剤及び記録材料
23日前
ダイキン工業株式会社
シラン化合物
16日前
花王株式会社
新規ピリジニウム化合物
22日前
株式会社トクヤマ
グルコンアミド誘導体の製造方法
16日前
株式会社トクヤマ
結晶形Iのリオシグアトの製造方法
1か月前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
1か月前
国立大学法人京都大学
細胞質送達ペプチド
13日前
ダイキン工業株式会社
分離方法
1か月前
石原産業株式会社
シアノイミダゾール系化合物の製造方法
1か月前
株式会社半導体エネルギー研究所
有機化合物、発光デバイス
1か月前
小川香料株式会社
化合物及び香料組成物
13日前
小川香料株式会社
化合物及び香料組成物
13日前
ダイキン工業株式会社
SF5含有シラン化合物
8日前
石原産業株式会社
シクラニリプロールの製造中間体の製造方法
1か月前
株式会社アイティー技研
炭化水素の合成方法及び合成装置
2か月前
オリザ油化株式会社
新規化合物及びその用途
1か月前
キッコーマン株式会社
ナノポアタンパク質
1か月前
株式会社レゾナック
C2化合物の製造方法
1か月前
続きを見る
他の特許を見る



 特許ウォッチ
特許ウォッチ