TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025038044
公報種別
公開特許公報(A)
公開日
2025-03-18
出願番号
2024216380,2023114162
出願日
2024-12-11,2018-05-08
発明の名称
炎症症状で使用するための抗IL-1R3抗体
出願人
サノフィ・バイオテクノロジー
代理人
個人
,
個人
主分類
C07K
16/24 20060101AFI20250311BHJP(有機化学)
要約
【課題】対象のIL1R3媒介炎症症状及び/又は障害の治療のための医薬の製造における、IL-1R3に特異的に結合する抗体又はその抗原結合断片の使用を提供する。
【解決手段】IL-1R3に特異的に結合する抗体又はその抗原結合断片は、特定のアミノ酸配列を含む重鎖可変(VH)領域と少なくとも95%同一であるVH領域及び特定のアミノ酸配列を含む軽鎖可変(VL)領域と少なくとも95%同一であるVL領域を含み、該抗体又はその抗原結合断片は、a)それぞれ特定のアミノ酸配列を含むCDR-H1、CDR-H2及びCDR-H3を含む相補性決定領域、及びb)それぞれ特定のアミノ酸配列を含むCDR-L1、CDR-L2及びCDR-L3を含む相補性決定領域、を含む、使用である。
【選択図】なし
特許請求の範囲
【請求項1】
対象のIL1R3媒介炎症症状及び/又は障害の治療における使用のための抗IL1R3抗体。
続きを表示(約 800 文字)
【請求項2】
前記炎症症状及び/又は障害が、炎症症状、免疫障害、線維性障害、好酸球性障害、感
染症、疼痛、中枢神経系障害、眼科障害、遺伝的全身性炎症疾患、及び全身性及び局所性の炎症疾患からなる群より選択される、請求項1に記載の抗IL1R3抗体。
【請求項3】
前記炎症症状が、高尿酸血症と関連する代謝性リウマチ障害である、請求項2に記載の抗IL1R3抗体。
【請求項4】
前記代謝性リウマチ障害が、痛風、偽痛風、薬物誘発性痛風及び慢性活動性(難治性)痛風の群から選択される、請求項3に記載の抗IL1R3抗体。
【請求項5】
前記IL1R3アンタゴニスト抗体が、痛風の治療のための治療薬と組み合わせて投与され
る、請求項4に記載の抗IL1R3抗体。
【請求項6】
前記炎症症状が、がん関連慢性炎症である、請求項2に記載の抗IL1R3抗体。
【請求項7】
前記IL1R3アンタゴニスト抗体が、1以上の細胞傷害性抗がん剤、細胞増殖抑制性抗が
ん剤及び/又は標的化抗がん剤と組み合わせて投与される、請求項6に記載の抗IL1R3抗体。
【請求項8】
前記対象が、1以上の細胞傷害性抗がん剤、細胞増殖抑制性抗がん剤及び/又は標的化
抗がん剤による治療に耐性があることを特徴とする、請求項6~7に記載の抗IL1R3抗体
。
【請求項9】
前記抗体のエフェクター機能が低下しているか、又は前記抗体のエフェクター機能が欠如している、請求項1~8のいずれか一項に記載の抗IL1R3抗体。
【請求項10】
前記抗体が、免疫細胞の枯渇を誘発しない、請求項1~9のいずれか一項に記載の抗IL1R3抗体。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、IL-1α、IL-1β、IL-33、IL-36、IL-RA、及び/又はIL1R3などのIL1R3シグナル伝達経路のメンバーの制御されない又は異常な発現を特徴とする医学的症状及び/又は
障害の治療方法に関する。より具体的には、本発明は、炎症症状及び/又は障害における
抗IL-1R3抗体の使用方法に関する。
続きを表示(約 13,000 文字)
【背景技術】
【0002】
インターロイキン1受容体アクセサリータンパク質(IL1RAP)(IL1R3とも称される)
は、1型インターロイキン1受容体(IL1R1)の共受容体であり、IL-1シグナル伝達の伝
達に不可欠である。IL-1の結合により、IL-1R1は、IL-1RAcPと会合して機能的シグナル伝達受容体複合体を形成し、その複合体がNFκB活性を刺激する。
【0003】
IL-33、その受容体ST2、及びIL-1RAcPも、IL-1β/IL-1R1/IL-1RAcP複合体としてNFκB
活性化に関して同様の活性を持つ複合体(IL-33/ST2/IL-1RAcP)を形成する。IL-36(IL-36α(IL-1F6)、IL-36β(IL-1F8)、及びIL-36γ(IL-1F9))、それらの受容体IL-36R、並びにIL-1RAcPもまた、IL-1β/IL-1R1/IL-1RAcP複合体としてNFκB活性に関して類似
の活性を有する複合体(IL-36/Il-36R/IL-1RAcP)を形成する。
【0004】
インターロイキン-1(IL-1)経路は、哺乳類の炎症反応で重要な役割を果たす細胞内シグナル伝達経路であり、広範囲の免疫学、代謝、生理学及び造血活動に関連する。IL-1ファミリーは、3つの構造的に関連するサイトカイン:IL-1アルファ、IL-1ベータ、及びIL-1受容体アンタゴニスト(IL-1ra)が含まれる。3つのうち、IL-1アルファ及びIL-1ベータは、炎症誘発性アゴニストであり、一方、IL-1受容体アンタゴニスト(IL-1ra)は、IL-1アルファ及びIL-1ベータ活性をブロックするよう機能する。IL-1のすべての既知の生物学的機能は、I型IL-1Rを通じて媒介される。IL-1アルファ、IL-1ベータ及びIL-1raは、I型IL-1Rと高い親和性で結合する。対照的に、IL-1ベータは高い親和性でII型IL-1Rに結合し
、IL-1アルファ及びIL-1raは低い親和性でII型IL-1Rに結合する。II型IL-1Rは、細胞質ドメインが大幅に切断されており、IL-1に結合すると細胞にシグナルを伝達しないが、代わりにデコイ受容体として作用することによりIL-1を媒介した反応の調節に関与する。
【0005】
IL-1の産生は、感染症、微生物毒素、炎症剤、アレルギー反応によって引き起こされる。IL-1の全体の主な機能は、炎症又はアレルギー性免疫反応の部位での免疫及び炎症反応の振幅と持続時間を調節することである。過剰なIL-1が生成される又はIL-1の発現が適切に調節されない場合、疾患状態が発症し得る。したがって、IL-1は様々な炎症性及び免疫調節性の疾患並びに症状に関係する。IL-1の全身的又は局所的な過剰が、多くの医学的障害の発生の一因となることが提案されている。この提案に加えて、IL-1アルファ及びIL-1ベータ活性をブロックするIL-1raは、IL-1シグナル伝達によって媒介されると考えられるいくつかの疾患の治療において様々な程度の効力を有することが示されている。
【0006】
IL-1の抑制は、異常若しくは過剰なIL-1発現又はIL-1活性により特徴づけられる様々な障害に苦しむ患者に有益であり得ることが示唆されている。IL-1ra及びICE阻害剤は、IL-1活性に関連する疾患の治療薬として限られた程度の成功しか収めていない。そのような
疾患の効果的な治療の考案は進歩しているが、改善された薬剤及び治療法が必要である。
【0007】
不幸にも、キネレット(Kineret)、イラリス(Ilaris)又はアルカリスト(Arcalyst
)などの既存のIL-1経路阻害剤には、それらの治療薬の使用を妨げる特定の欠点がある。例えば、キネレット(IL-1Ra)は半減期が短く、それゆえ非常に頻繁な治療間隔(毎日)
が必要である。単一のサイトカイン(イラリスなど)を標的とする抗体により、他のIL-1ファミリーサイトカインの不必要なシグナル伝達/活性が発生する。さらに、IL-1ファミ
リーのサイトカインは、炎症性及びIL-1R3非依存性抗炎症シグナル伝達の両方を誘発することが知られており、いずれもサイトカイン又はそれらのアルファ鎖受容体を標的とする場合に干渉される。対照的に、本発明の抗体は、治療間隔の低減、異なるIL-1ファミリーサイトカイン媒介シグナル伝達の同時阻害、及び炎症性シグナル伝達経路のブロックに関する特異性という利点を兼ね備えている。
【発明の概要】
【0008】
したがって、本発明は、以下に記載される好ましい特徴を有するIL1R3シグナル伝達経
路のメンバーの制御されない発現に関連する症状を治療するための新規及び改善された方法を提供する。
【図面の簡単な説明】
【0009】
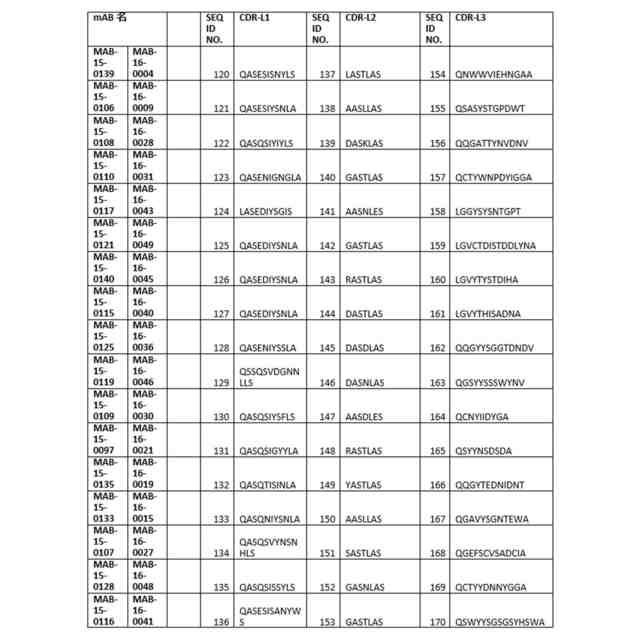
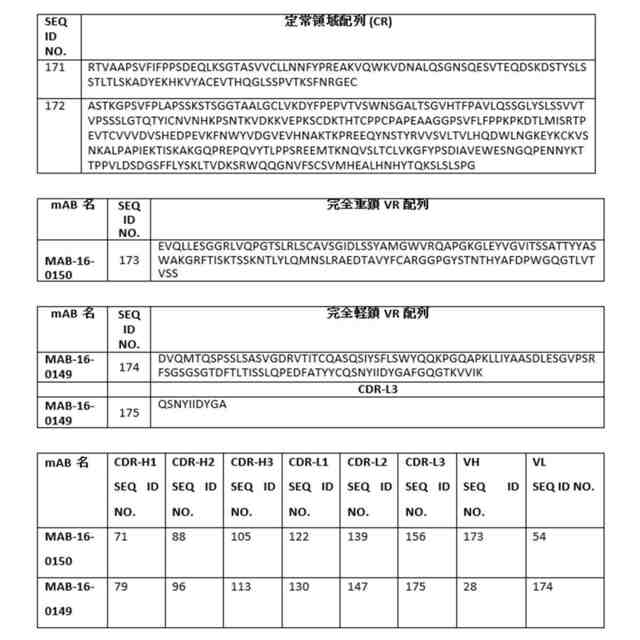
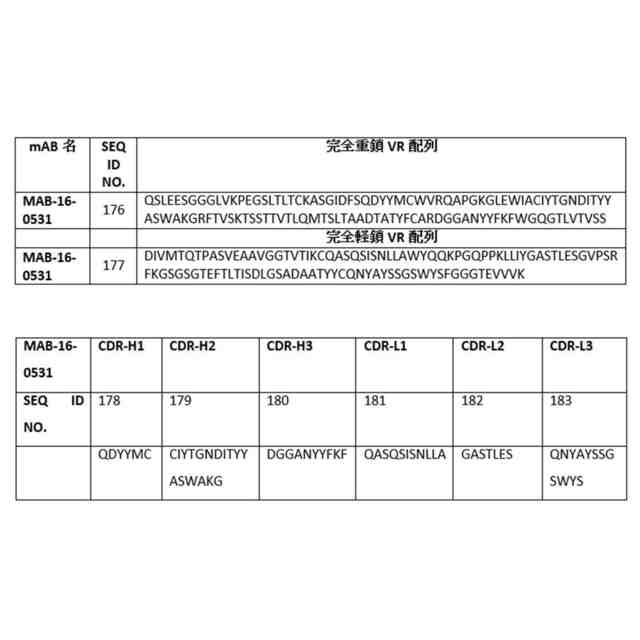
配列(一文字コードのアミノ酸) 可変領域(VR)の完全配列: 重鎖:VH完全:配列番号1~34並びに配列番号173及び176 軽鎖:VL完全:配列番号35~68並びに配列番号174及び177 相補性決定領域(CDR): 重鎖:CDRH1:配列番号69~85及び178CDRH2:配列番号86~102及び179CDRH3:配列番号103~119及び180 軽鎖:CDRL1:配列番号120~136及び181CDRL2:配列番号137~153及び182CDRL3:配列番号154~170及び175及び183 定常領域(CR): 軽鎖:CR-L:配列番号171重鎖:CR-H:配列番号172 次の図では、AF676は次のリンク:https//www.rndsystems.com/products/human-il-1-racp-il-1-r3-antibody_af676から購入した市販のポリクローナル抗体調整物である。
配列(一文字コードのアミノ酸) 可変領域(VR)の完全配列: 重鎖:VH完全:配列番号1~34並びに配列番号173及び176 軽鎖:VL完全:配列番号35~68並びに配列番号174及び177 相補性決定領域(CDR): 重鎖:CDRH1:配列番号69~85及び178CDRH2:配列番号86~102及び179CDRH3:配列番号103~119及び180 軽鎖:CDRL1:配列番号120~136及び181CDRL2:配列番号137~153及び182CDRL3:配列番号154~170及び175及び183 定常領域(CR): 軽鎖:CR-L:配列番号171重鎖:CR-H:配列番号172 次の図では、AF676は次のリンク:https//www.rndsystems.com/products/human-il-1-racp-il-1-r3-antibody_af676から購入した市販のポリクローナル抗体調整物である。
配列(一文字コードのアミノ酸) 可変領域(VR)の完全配列: 重鎖:VH完全:配列番号1~34並びに配列番号173及び176 軽鎖:VL完全:配列番号35~68並びに配列番号174及び177 相補性決定領域(CDR): 重鎖:CDRH1:配列番号69~85及び178CDRH2:配列番号86~102及び179CDRH3:配列番号103~119及び180 軽鎖:CDRL1:配列番号120~136及び181CDRL2:配列番号137~153及び182CDRL3:配列番号154~170及び175及び183 定常領域(CR): 軽鎖:CR-L:配列番号171重鎖:CR-H:配列番号172 次の図では、AF676は次のリンク:https//www.rndsystems.com/products/human-il-1-racp-il-1-r3-antibody_af676から購入した市販のポリクローナル抗体調整物である。
配列(一文字コードのアミノ酸) 可変領域(VR)の完全配列: 重鎖:VH完全:配列番号1~34並びに配列番号173及び176 軽鎖:VL完全:配列番号35~68並びに配列番号174及び177 相補性決定領域(CDR): 重鎖:CDRH1:配列番号69~85及び178CDRH2:配列番号86~102及び179CDRH3:配列番号103~119及び180 軽鎖:CDRL1:配列番号120~136及び181CDRL2:配列番号137~153及び182CDRL3:配列番号154~170及び175及び183 定常領域(CR): 軽鎖:CR-L:配列番号171重鎖:CR-H:配列番号172 次の図では、AF676は次のリンク:https//www.rndsystems.com/products/human-il-1-racp-il-1-r3-antibody_af676から購入した市販のポリクローナル抗体調整物である。
配列(一文字コードのアミノ酸) 可変領域(VR)の完全配列: 重鎖:VH完全:配列番号1~34並びに配列番号173及び176 軽鎖:VL完全:配列番号35~68並びに配列番号174及び177 相補性決定領域(CDR): 重鎖:CDRH1:配列番号69~85及び178CDRH2:配列番号86~102及び179CDRH3:配列番号103~119及び180 軽鎖:CDRL1:配列番号120~136及び181CDRL2:配列番号137~153及び182CDRL3:配列番号154~170及び175及び183 定常領域(CR): 軽鎖:CR-L:配列番号171重鎖:CR-H:配列番号172 次の図では、AF676は次のリンク:https//www.rndsystems.com/products/human-il-1-racp-il-1-r3-antibody_af676から購入した市販のポリクローナル抗体調整物である。
配列(一文字コードのアミノ酸) 可変領域(VR)の完全配列: 重鎖:VH完全:配列番号1~34並びに配列番号173及び176 軽鎖:VL完全:配列番号35~68並びに配列番号174及び177 相補性決定領域(CDR): 重鎖:CDRH1:配列番号69~85及び178CDRH2:配列番号86~102及び179CDRH3:配列番号103~119及び180 軽鎖:CDRL1:配列番号120~136及び181CDRL2:配列番号137~153及び182CDRL3:配列番号154~170及び175及び183 定常領域(CR): 軽鎖:CR-L:配列番号171重鎖:CR-H:配列番号172 次の図では、AF676は次のリンク:https//www.rndsystems.com/products/human-il-1-racp-il-1-r3-antibody_af676から購入した市販のポリクローナル抗体調整物である。
配列(一文字コードのアミノ酸) 可変領域(VR)の完全配列: 重鎖:VH完全:配列番号1~34並びに配列番号173及び176 軽鎖:VL完全:配列番号35~68並びに配列番号174及び177 相補性決定領域(CDR): 重鎖:CDRH1:配列番号69~85及び178CDRH2:配列番号86~102及び179CDRH3:配列番号103~119及び180 軽鎖:CDRL1:配列番号120~136及び181CDRL2:配列番号137~153及び182CDRL3:配列番号154~170及び175及び183 定常領域(CR): 軽鎖:CR-L:配列番号171重鎖:CR-H:配列番号172 次の図では、AF676は次のリンク:https//www.rndsystems.com/products/human-il-1-racp-il-1-r3-antibody_af676から購入した市販のポリクローナル抗体調整物である。
配列(一文字コードのアミノ酸) 可変領域(VR)の完全配列: 重鎖:VH完全:配列番号1~34並びに配列番号173及び176 軽鎖:VL完全:配列番号35~68並びに配列番号174及び177 相補性決定領域(CDR): 重鎖:CDRH1:配列番号69~85及び178CDRH2:配列番号86~102及び179CDRH3:配列番号103~119及び180 軽鎖:CDRL1:配列番号120~136及び181CDRL2:配列番号137~153及び182CDRL3:配列番号154~170及び175及び183 定常領域(CR): 軽鎖:CR-L:配列番号171重鎖:CR-H:配列番号172 次の図では、AF676は次のリンク:https//www.rndsystems.com/products/human-il-1-racp-il-1-r3-antibody_af676から購入した市販のポリクローナル抗体調整物である。
ヒトIL-1R3 ELISA 384-マイクロタイタープレートを、IL-1R3のヒト細胞外ドメインを表すヒトIL-1R3タンパク質でコーティングした(0.5mg/ml、少なくとも1時間)。集中的な洗浄ステップとそれに続くブロッキングステップの後、抗体を加え(ウェルあたり12,5μl)及び室温で1時間インキュベートした。非結合抗体は集中的に洗い流した。結合した抗体の量は、マイクロタイタープレートでペルオキシダーゼ標識抗ヒト検出抗体とインキュベートすることにより同定した(室温で1時間)。TMBを添加によりペルオキシダーゼ反応を開始し、450nm/620nmで吸光度を測定した。
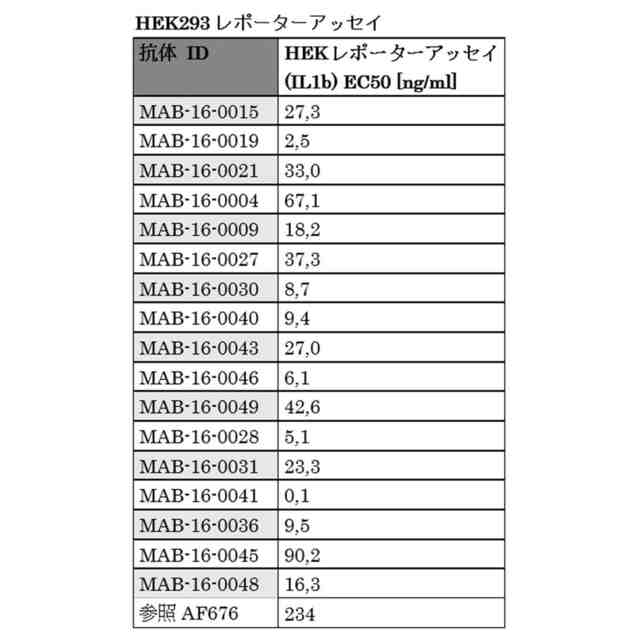
HEK293レポーターアッセイ HEK293T/17-FR細胞にpGL4.32[luc2P/NF-κB-RE/Hygro]ベクター(Promega)を安定的にトランスフェクトし、384ウェルPDL Costar細胞培養プレートに播種し、その後抗体とともに30分間インキュベートした。次に、細胞をIL-1βで5時間刺激し、その後製造業者のプロトコルにより、Steady-Glo Luciferase アッセイキット(Promega)を使用してNF-kB活性を測定した。
A549安定発現株を使用したNFκBルシフェラーゼレポーターアッセイ A549-NFkB-RE-Luc安定トランスフェクト細胞(Signosisから購入)を3日間培養した(1,7E+04 細胞/cm
3
)。384ウェルの低フランジ白色平底ポリスチレンTC処理マイクロタイタープレート(Corning)にウェルあたり4x10
4
個の細胞を充填した。10時間の培養期間の後、細胞を抗体とともに1時間インキュベートし、その後細胞を10μlのIL-1βでさらに5時間刺激した。NFkB調節は、Steady-Glo(商標) ルシフェラーゼアッセイシステム(Promega)によって測定され、非刺激細胞に関連する各ウェルの相対発光単位を決定する。
細胞結合分析:IL-1R3発現細胞への結合 ヒト化抗IL-1R3 IgG1-LALA抗体は、フローサイトメトリーを使用して、異なるIL-1R3受容体密度の細胞株への結合についてテストした。ヒト化抗IL-1R3 IgG1-LALA抗体は、低及び高IL-1R3発現細胞株に結合する。抗体はマウスNIH-3T3細胞には結合しない。実験は、実施例4に記載の方法により実施した。
細胞結合分析:ヒトIL-1R3高発現細胞株SK-MEL-30への細胞結合 ヒト化抗IL-1R3 IgG1-LALA抗体のEC50細胞結合値は、フローサイトメトリーを使用して高IL-1R3発現細胞株SK-MEL-30に結合することにより決定された。ヒト化抗IL-1R3 IgG1-LALA抗体MAB-16-0030及びMAB-16-0149は、それぞれ307及び306 ng/mlの細胞結合を示す。実験は、実施例4に記載の方法により実施した。
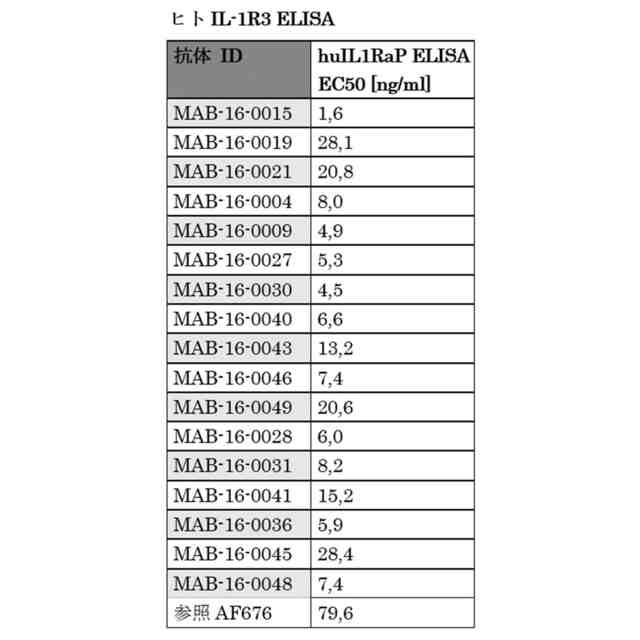
ヒト-IL-1R3生化学的ELISA 組換えヒトIL-1R3タンパク質へのヒト化抗IL-1R3 IgG1-LALA抗体の結合を生化学的ELISAでテストした。例示された抗体は、それぞれ16,3ng/ml及び29,1ng/mlのEC50結合値を示す。実験は、実施例5に記載の方法により実施した。
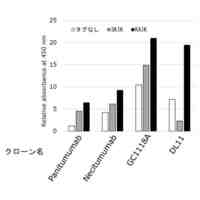
A549-NFkB-RE-Luc細胞におけるNfKBシグナル伝達を媒介したヒトIL-1a及びIL-1bの阻害 IL-1a及びIL-1bの機能的中和は、それぞれ0.1ng/mlのIL-1a及びIL-1bで刺激したA549-NFkB-RE-Luc細胞を用いた細胞ベースの遺伝子レポーターアッセイでテストした。ヒト化抗IL-1R3 IgG1-LALA抗体は、ヤギ抗ヒトIL1-R3ポリクローナル抗体AF676(R&D Systems)よりも優れたEC50値を示す。実験は、実施例6に記載の方法によって実施した。
IL-1α及びIL-1β機能的中和アッセイ - A-549細胞によるヒトIL-1a及びIL-1b媒介IL-6放出の阻害 ヒト化抗IL-1R3 IgG1-LALA抗体によるIL-6のIL-1a及びIL-1b媒介細胞放出の中和は、A-549細胞を使用してテストされた。EC50値は、ヒト化抗IL-1R3 IgG1-LALA抗体がヤギ抗ヒトIL1-R3ポリクローナル抗体AF676(R&D Systems)よりも優れていることを示す。実験は、実施例7に記載の方法によって実施した。
IL-33機能的中和アッセイ - HEK-Blue-IL33
TM
細胞でのヒトIL-33を媒介したNfkBシグナル伝達の阻害 ヒト化抗IL-1R3 IgG1-LALA抗体によるIL-33媒介細胞シグナル伝達の中和を、IL-33刺激遺伝子レポーターHEK-Blue-IL33
TM
細胞(InvivoGen)を使用して、テストした。EC50値は、ヒト化抗IL-1R3 IgG1-LALA抗体がヤギ抗ヒトIL1-R3ポリクローナル抗体AF676(R&D Systems)よりも優れていることを示す。実験は、実施例8に記載の方法により実施した。
IL-36機能的中和アッセイ - HEK-293/17-IF細胞におけるヒトIL-36を媒介したNfkBシグナル伝達の阻害 ヒト化抗IL-1R3 IgG1-LALA抗体によるIL-36媒介細胞シグナル伝達の中和を、IL-36g刺激遺伝子レポーターHEK-293/17-IF細胞を使用してテストされた。典型的なヒト化抗IL-1R3 IgG1-LALA抗体は、ヤギ抗ヒトIL1-R3ポリクローナル抗体AF676(R&D Systems)よりも優れたEC50値を示す。実験は、実施例9に記載の方法により実施した。
IL-1a、IL-33、及びIL-36aを媒介した細胞性サイトカイン放出の中和 IL-1a、IL-33、及びIL-36aを媒介した細胞性サイトカイン放出の中和は、特定のIL-1a、IL-33、及びIL-36a依存性細胞システムを使用してテストされた。本発明による代表的なヒト化抗IL-1R3 IgG1-LALA抗体(MAB-16-0030)によるサイトカイン放出の阻害をテストし、及びIL-1Raと比較した。本発明による抗体は3つすべての刺激により媒介されるサイトカイン放出を阻害することができたが、IL-1RaはIL-1a媒介サイトカイン放出のみに影響した。実験は、実施例10に記載の方法により実施した。
ヒト化抗IL-1R3 IgG1-LALA抗体で処理した非刺激PBMCの生存率とIL-6放出 免疫細胞への抗体の結合は、細胞の枯渇及び有害な影響を、例えばアポトーシスシグナル伝達経路の直接誘導、過剰なサイトカイン放出の刺激、又は抗体依存性細胞傷害(ADCC)によって、もたらし得る。ヒト化抗IL-1R3 IgG1-LALA抗体がPBMCの生存率に直接影響することを除外するために、3つのドナーのPBMCの生存率及びIL-6放出を、本発明による代表的なヒト化抗IL-1R3 IgG1- LALA抗体(MAB-16-0030)の異なる濃度で、1、3及び5日インキュベート後、調査した。生存率もIL-6放出も影響を受けなかった。これらの結果は、ヒト化抗IL-1R3 IgG1-LALA抗体が、状況に応じた細胞の枯渇及び細胞に有害な影響の誘導なしに、免疫細胞のIL-1R3機能をブロックすることを支持する。実験は、実施例11に記載の方法により実施した。
ヒト化抗IL-1R3 IgG1-LALA抗体で処理した非刺激PBMCの生存率とIL-6放出 免疫細胞への抗体の結合は、細胞の枯渇及び有害な影響を、例えばアポトーシスシグナル伝達経路の直接誘導、過剰なサイトカイン放出の刺激、又は抗体依存性細胞傷害(ADCC)によって、もたらし得る。ヒト化抗IL-1R3 IgG1-LALA抗体がPBMCの生存率に直接影響することを除外するために、3つのドナーのPBMCの生存率及びIL-6放出を、本発明による代表的なヒト化抗IL-1R3 IgG1- LALA抗体(MAB-16-0030)の異なる濃度で、1、3及び5日インキュベート後、調査した。生存率もIL-6放出も影響を受けなかった。これらの結果は、ヒト化抗IL-1R3 IgG1-LALA抗体が、状況に応じた細胞の枯渇及び細胞に有害な影響の誘導なしに、免疫細胞のIL-1R3機能をブロックすることを支持する。実験は、実施例11に記載の方法により実施した。
異なる刺激で活性化されたPBMCの機能的ブロック ヒト化抗IL-1R3 IgG1-LALA抗体が特定又は複合的な刺激で刺激されたPBMCの活性を阻害するかどうかをテストするために、10体のドナーのPBMCをLPS、熱不活性化カンジダアルビカンス、IL-12/IL-33又は抗CD3/CD28抗体で刺激した。本発明の代表的なヒト化抗IL-1R3 IgG1-LALA抗体(MAB-16-0030)は、テストしたすべての刺激によって媒介されるサイトカイン放出を阻害することができた。実験は、実施例12に記載の方法により実施した。
異なる刺激で活性化されたPBMCの機能的ブロック ヒト化抗IL-1R3 IgG1-LALA抗体が特定又は複合的な刺激で刺激されたPBMCの活性を阻害するかどうかをテストするために、10体のドナーのPBMCをLPS、熱不活性化カンジダアルビカンス、IL-12/IL-33又は抗CD3/CD28抗体で刺激した。本発明の代表的なヒト化抗IL-1R3 IgG1-LALA抗体(MAB-16-0030)は、テストしたすべての刺激によって媒介されるサイトカイン放出を阻害することができた。実験は、実施例12に記載の方法により実施した。
カンジダアルビカンスで活性化された全血中の免疫細胞の機能的ブロック ヒト化抗IL-1R3 IgG1-LALA抗体が全血中の免疫細胞の活性化を阻害するかどうかをテストするために、8体のドナーの全血を熱不活化カンジダアルビカンスで刺激した。本発明による代表的なヒト化抗IL-1R3 IgG1-LALA抗体(MAB-16-0030)は、カンジダ誘発IL-6サイトカイン放出を阻害することができた。実験は、実施例13に記載の方法により実施した。
混合リンパ球反応(MLR)におけるサイトカイン放出のブロック ヒト化抗IL-1R3 IgG1-LALA抗体が多様なサイトカインの放出をブロックする能力を、健康で一致しない(unmatched)ドナーのPBMCを使用した混合リンパ球反応(MLR)でテストした。本発明による代表的なヒト化抗IL-1R3 IgG1-LALA抗体(MAB-16-0030)は、IFNg、IL-6、TNF-a、IL-13、IL-17及びIL-10の放出を阻害することができた。実験は、実施例14に記載の方法により実施した。
混合リンパ球反応(MLR)におけるサイトカイン放出のブロック ヒト化抗IL-1R3 IgG1-LALA抗体が多様なサイトカインの放出をブロックする能力を、健康で一致しない(unmatched)ドナーのPBMCを使用した混合リンパ球反応(MLR)でテストした。本発明による代表的なヒト化抗IL-1R3 IgG1-LALA抗体(MAB-16-0030)は、IFNg、IL-6、TNF-a、IL-13、IL-17及びIL-10の放出を阻害することができた。実験は、実施例14に記載の方法により実施した。
混合リンパ球反応(MLR)におけるサイトカイン放出のブロック ヒト化抗IL-1R3 IgG1-LALA抗体が多様なサイトカインの放出をブロックする能力を、健康で一致しない(unmatched)ドナーのPBMCを使用した混合リンパ球反応(MLR)でテストした。本発明による代表的なヒト化抗IL-1R3 IgG1-LALA抗体(MAB-16-0030)は、IFNg、IL-6、TNF-a、IL-13、IL-17及びIL-10の放出を阻害することができた。実験は、実施例14に記載の方法により実施した。
ネズミ科(murine)の細胞におけるIL-1b媒介シグナル伝達の中和 本発明による抗マウスIL-1R3抗体によるIL-1bの機能的中和を、NFkBルシフェラーゼ遺伝子レポーターアッセイに基づくマウスNIH-3T3細胞を使用してテストした。本発明による代表的な抗体(MAB-16-0531)は、805ng/mlのEC50でIL-1b媒介NFkBシグナル伝達の用量依存性阻害を示した。実験は、実施例15に記載の方法により実施した。
ネズミ科(murine)の細胞からのIL-1b媒介IL-6放出の中和 本発明による抗マウスIL-1R3抗体によるマウス細胞からのIL-6放出の阻害を、IL-1b刺激NIH-3T3細胞でテストした。本発明による代表的な抗体(MAB-16-0531)は、1560ng/mlのEC50でIL-6放出の用量依存性阻害を示した。実験は、実施例16に記載の方法により実施した。
尿酸一ナトリウム結晶(MSU)誘発マウス腹膜炎モデル 痛風患者に抗IL-R3療法を適用する理論的根拠をテストするために、尿酸一ナトリウム結晶(MSU)誘発マウス腹膜炎モデルが使用された。MSUの投与は、腹膜腔に強い炎症反応を産生し、いくつかの炎症細胞(単球及び好中球など)の流入を誘導し、急性痛風反応の古典的な炎症プロファイルを模倣する。本発明による代表的な抗IL1R3抗体(MAB-16-0531)又はIL-1Raを、腹膜に3mg MSUを注射する1時間前にIP投与した。腹膜腔内の細胞の測定は、抗IL-1R3抗体治療がリンパ球、単球及び顆粒球のMSU誘導浸潤を有意に阻害することを示している。実験は、実施例17に記載の方法により実施した。
MSU腹膜炎における好中球の活性及びサイトカイン産生 MSU腹膜炎における炎症の局所及び全身マーカーを含む好中球活性の衰弱。(a)IP液中の好中球プロテアーゼエラスターゼのレベル。(b)IP液からの細胞における細胞内エラスターゼの合計レベル。(c)IP液からの細胞における細胞内MPOの合計レベル。(d)局所的(IP液体)及び全身的なサイト-及びケモカイン濃度。WB; 溶解した全血、標準(norm.);脾臓中のタンパク質のmgあたりの標準化。MSU;ビヒクル+MSU。a-mIL1R3;a-mIL1R3+MSU。IL 1Ra;IL 1Ra+MSU。MSU刺激群と比較した阻害剤処理群。
*
p<0.05、
**
p<0.01、
***
p<0.001、
****
p<0.0001(スチューデントt検定、平均+SEM、除く:(b)、G CSF、KC(WB)及びKC(脾臓);マン・ホイットニーU検定、IQRの中央値)。
OVAアレルギー性喘息のin vivoモデル a-mIL1R3抗体は、in vivoでのOVA誘発細胞流入を減少させる。(a)BAL液からの総WBC。(b)フロー規定細胞は、(a)とは異なる。(c)H&E及びPAS染色肺組織を使用したグループ平均(WBC)の太いドットマークされたマウスの組織構造。OVA刺激+a-mIL1R3(MAB-16-0531)処理マウス及びOVA単独との比較。
*
p<0.05、
**
p<0.01、
***
p<0.001(スチューデントt検定)。平均+SEM。
OVAアレルギー性喘息のin vivoモデル a-mIL1R3抗体は、in vivoでのOVA誘発細胞流入を減少させる。(a)BAL液からの総WBC。(b)フロー規定細胞は、(a)とは異なる。(c)H&E及びPAS染色肺組織を使用したグループ平均(WBC)の太いドットマークされたマウスの組織構造。OVA刺激+a-mIL1R3(MAB-16-0531)処理マウス及びOVA単独との比較。
*
p<0.05、
**
p<0.01、
***
p<0.001(スチューデントt検定)。平均+SEM。
in vivoモデルのイミキモド乾癬 (a)皮膚におけるIL36αmRNAの誘導。(b)IMQ及びコントロールクリームの間で適用された耳の厚さの倍率変化。(c)研究終了時にn=6のブラインドによる視覚スコアリング(0;影響なし-4;最大の影響(紅斑及びスケーリング))。(d)タンパク質のmgあたりで標準化された皮膚のMPO濃度。(e)(c)及び(d)からのデータ間のスピアマン相関。(e)全血中の顆粒球濃度。阻害剤処理及びIMQ刺激群間の比較。
*
p<0.05、
**
p<0.01、
***
p<0.001、
****
p<0.0001、((b、f)マン・ホイットニーU検定(d、e)スチューデント検定、平均±SEM(d)IQRの中央値を除く)。
in vivoモデルのイミキモド乾癬 (a)皮膚におけるIL36αmRNAの誘導。(b)IMQ及びコントロールクリームの間で適用された耳の厚さの倍率変化。(c)研究終了時にn=6のブラインドによる視覚スコアリング(0;影響なし-4;最大の影響(紅斑及びスケーリング))。(d)タンパク質のmgあたりで標準化された皮膚のMPO濃度。(e)(c)及び(d)からのデータ間のスピアマン相関。(e)全血中の顆粒球濃度。阻害剤処理及びIMQ刺激群間の比較。
*
p<0.05、
**
p<0.01、
***
p<0.001、
****
p<0.0001、((b、f)マン・ホイットニーU検定(d、e)スチューデント検定、平均±SEM(d)IQRの中央値を除く)。
in vivoモデルのイミキモド乾癬 イミキモド処理マウスの皮膚生検におけるIL36α/β/γ、IL17F及びTNFa mRNAレベル
in vivoモデルのイミキモド乾癬 イミキモド処理マウスの皮膚生検におけるIL36α/β/γ、IL17F及びTNFa mRNAレベル
抗IL1R3 IgG1及びIgG1-LALA抗体のエフェクター細胞媒介機能 IgG1-LALA又はIgG1 Fc部分のいずれかを含む、抗IL1R3抗体MAB-16-0030を、エフェクター細胞におけるFc受容体媒介NF-kBシグナル伝達の活性についてテストした。増加する濃度のIgG1抗体のhIL1R3発現SK-MEL-30細胞への結合は、IgG1-LALA抗体とは対照的に、Jurkat遺伝子レポーターエフェクター細胞におけるFc受容体媒介シグナル伝達を誘発する。
【発明を実施するための形態】
【0010】
本発明は、IL-1α、IL-1β、IL-33、IL-36、IL1RA及び/又はIL1R3、同様にそのバリア
ントなどのIL1R3シグナル伝達経路のメンバーの制御されない又は異常な発現によって特
徴づけられる医学的症状及び/又は障害の治療のための方法に関する。
(【0011】以降は省略されています)
この特許をJ-PlatPat(特許庁公式サイト)で参照する
関連特許
東ソー株式会社
タンパク質の発現方法
17日前
株式会社トクヤマ
四塩化炭素の製造方法
2か月前
東ソー株式会社
炭素-窒素結合形成方法
2か月前
株式会社トクヤマ
シロキサン類の回収方法
2か月前
株式会社トクヤマ
ビオチン誘導体の製造方法
2か月前
株式会社半導体エネルギー研究所
有機化合物
1か月前
株式会社トクヤマ
ベンザゼピン化合物の製造方法
1か月前
株式会社コスモス
液状炭化水素の増産方法
2か月前
株式会社コスモス
液状炭化水素の増産方法
2か月前
株式会社トクヤマ
ホルムアミド化合物の製造方法
1か月前
株式会社トクヤマ
チオラクトン誘導体の製造方法
7日前
東ソー株式会社
イソシアネート化合物の製造方法
1か月前
日本特殊陶業株式会社
メタン製造装置
3か月前
ユニチカ株式会社
ビスマレイミドおよびその製造方法
4か月前
artience株式会社
四塩基酸無水物の製造方法
3か月前
信越化学工業株式会社
新規化合物
3か月前
金剛化学株式会社
ボルチオキセチンの製造方法
4か月前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
2か月前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
3か月前
大阪瓦斯株式会社
メタン製造システム
3日前
株式会社トクヤマ
サフィナミド若しくはその塩の製造方法
3か月前
本田技研工業株式会社
CO2変換方法
3日前
三菱ケミカル株式会社
アルコールの製造方法
25日前
国立大学法人東京農工大学
深共晶溶媒
1か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
17日前
東ソー株式会社
免疫グロブリン結合性タンパク質の保存溶液
3か月前
JNC株式会社
有機ケイ素化合物およびこれを用いた重合体
2か月前
キヤノン株式会社
有機化合物及び有機発光素子
4か月前
キヤノン株式会社
有機化合物及び有機発光素子
4か月前
株式会社トクヤマ
アシル化ベンゼン誘導体の酸塩の製造方法
1か月前
キヤノン株式会社
有機化合物及び有機発光素子
7日前
キヤノン株式会社
有機化合物及び有機発光素子
3か月前
JNC株式会社
有機ケイ素化合物およびこれを用いた重合体
1か月前
旭化成株式会社
トリオキサンの製造方法
3か月前
株式会社半導体エネルギー研究所
有機金属錯体、発光デバイス
4か月前
株式会社半導体エネルギー研究所
有機金属錯体、発光デバイス
1か月前
続きを見る
他の特許を見る




 特許ウォッチ
特許ウォッチ