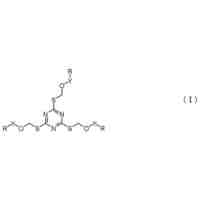
発明の詳細な説明【技術分野】 【0001】 本願の発明は、ヒト抗IL-33中和モノクローナル抗体及び当該抗体に競合する抗体、並びにこれらの抗体を含むサイトカイン発現抑制剤、及びIL-33関連疾患の治療、予防または軽減用医薬組成物に関する。 続きを表示(約 5,300 文字)【背景技術】 【0002】 インターロイキン-33(IL-33)は、炎症性状態において役割を果たすと考えられているインターロイキン-1ファミリーに属するサイトカインである。IL-33は、上皮細胞や血管内皮細胞の核内で恒常的に発現しており、感染や物理的・化学的ストレスによる組織傷害によって細胞破壊と共に放出され、アラーミンとして機能する。また、IL-33の発現はリポ多糖等の刺激によって上昇し、分泌される機構もあると考えられている。細胞外に放出されたIL-33は、細胞上に発現するIL-33受容体に結合することによって、細胞内シグナルを活性化することができる。IL-33受容体は、様々な免疫系細胞や上皮細胞などで発現しており、これらの細胞において、IL-33誘導性の細胞内シグナル伝達が生じる。 【0003】 IL-33は、IL-33受容体を発現する免疫系細胞の内、Th2細胞、マスト細胞、好酸球、好塩基球、NK(natural killer)T細胞やグループ2自然リンパ球からのTh2サイトカイン(IL-4、IL-5、IL-6、IL-13等)の産生を誘導することによって、アレルギー性炎症(喘息、アトピー性皮膚炎、花粉症、アナフィラキシーショックなど)を誘導すると考えられている(非特許文献1:Tatsukuni Ohno et al., Allergy, 2012, Vol. 67, p1203)。また、IL-33受容体を発現する免疫系細胞のうちマスト細胞やマクロファージでは、IL-33刺激によりIL-1β、IL-6、TNF(tumor necrosis factor)-αの産生が誘導され、これが自己抗体誘導関節炎(関節リウマチのモデル)の発症に関与していることが示唆されている(非特許文献2:Damo Xu et al., Journal of Immunology, 2010, Vol. 184, p2620)。IL-33のアンタゴニストが急性腎障害に有効であることが示唆されている(非特許文献3:Ali Akcay et al., Journal of American Society Nephrology, 2011, Vol.22, p2057) 。ヒトにおいては、IL-33の発現上昇が、種々の炎症性疾患(関節リウマチ、喘息、全身性硬化症、肝線維症、肺線維症等の線維症、乾癬、潰瘍性大腸炎、クローン病、多発性硬化症、強直性脊椎炎等)で認められており、IL-33が様々な疾患の発症・維持に関与していると考えられている(非特許文献4:Yasushi Matsuyama et al., Journal of Rheumatology, 2010, Vol. 37, p18;非特許文献5:David Pre´fontaine et al., Journal of Allergy and Clinical Immunology, 2010, Vol. 125, p752;非特許文献6:Koichi Yanaba et al., Clinical Rheumatology, 2011, Vol. 30, p825;非特許文献7:A. L. Rankin et al., Journal of Immunology, 2010, Vol. 184, p1526;非特許文献8:Tamar Mchedlidze et al., Immunity, 2013, Vol. 39, p357;非特許文献9:Liang-An Hu et al., Asian Pacific Journal of Cancer Prevention, 2013, Vol. 14, p2563;非特許文献10:Luca Pastorelli et al., Proceedings of the National Academy of Sciences of the United States of America, 2010, vol. 107, p8017)。 【0004】 このように様々な疾患、特に炎症性疾患におけるIL-33の関与が知られていることから、IL-33のアゴニストやアンタゴニストの開発が行われてきている(特許文献1から4)。なかでも、その特異性や作用強度の点からIL-33に対する抗体が注目されてきている。しかしながら、これまで開発されてきた抗体は、エピトープの特定がないマウス抗体(特許文献1)、またはカスパーゼによるIL-33の切断部位を特定し、活性型が非切断IL-33であることを特定したことに基づき、かかる切断部位を含む領域(配列表の配列番号226の残基155から残基198)をエピトープとした抗体(特許文献2)や市販のヤギポリクローナル抗体が知られていた。2014年1月10日にAnaptysBio社が同社のホームページにて独自の体細胞超変異技術(SHM―XEL)プラットフォームを用いてIL-33に対する治療用開発候補抗体ANB020の作製に成功したことを報告した(非特許文献11:Hamza Suria, ’AnaptysBio announces development of novel anti-IL-33 therapeutic antibody’, [on line], 2014, [retrieved on 11 January 2014], Retrieved from Internet:<URL: http://www.anaptysbio.com/anti-il-33/>) 。また、Murphyらはヒト抗体の可変領域遺伝子を導入したマウスであるVelocImmuneマウスを用いて20種類のヒト抗IL-33モノクローナル抗体を取得した(特許文献5)。しかしながら、該抗体のエピトープは開示されておらず、また20種類のヒト抗IL-33モノクローナル抗体のフレームワーク領域のアミノ酸配列は2アミノ酸残基以上でヒト生殖系列のアミノ酸配列と異なっている。したがって、これらの抗体をヒトに投与した場合に、当該抗体に対する免疫反応が惹起され、ヒト抗ヒト免疫グロブリン抗体(HAHA)の誘導により、作用効果の低減、炎症やその他の副作用の誘発という問題点がある。 【先行技術文献】 【特許文献】 【0005】 国際公開第2005/079844号 国際公開第2008/132709号 国際公開第2011/031600号 国際公開第2008/144610号 国際公開第2014/164959号 【非特許文献】 【0006】 Tatsukuni Ohno et al., Allergy, 2012, Vol. 67, p120 Damo Xu et al., Journal of Immunology, 2010, Vol. 184, p2620 Ali Akcay et al., Journal of American Society Nephrology, 2011, Vol.22, p2057 Yasushi Matsuyama et al., Journal of Rheumatology, 2010, Vol. 37, p18 David Pre´fontaine et al., Journal of Allergy and Clinical Immunology, 2010, Vol. 125, p752 Koichi Yanaba et al., Clinical Rheumatology, 2011, Vol. 30, p825 A. L. Rankin et al., Journal of Immunology, 2010, Vol. 184, p1526 Tamar Mchedlidze et al., Immunity, 2013, Vol. 39, p357 Liang-An Hu et al., Asian Pacific Journal of Cancer Prevention, 2013, Vol. 14, p2563 Luca Pastorelli et al., Proceedings of the National Academy of Sciences of the United States of America, 2010, vol. 107, p8017 Hamza Suria, ’AnaptysBio announces development of novel anti―IL―33 therapeutic antibody‘, [on line], 2014, [retrieved on 11 January 2014], Retrieved from Internet:<URL: http://www.anaptysbio.com/anti―il―33/> 【発明の概要】 【発明が解決しようとする課題】 【0007】 近年、IL-33と疾患との関係が明らかになってきており、IL-33のアンタゴニスト作用を有する抗IL-33中和モノクローナル抗体が望まれている。抗IL-33中和モノクローナル抗体の作用は、その抗体が結合するエピトープの領域と密接に関わっている。IL-33は細胞破壊とともに細胞外へ放出されるため、リソゾーム由来のタンパク質分解酵素等により切断される可能性が高く、いわゆる成熟型IL-33、さらに成熟型IL-33から派生する、IL-33の活性を有する多くの断片が生じ得る。IL-33の連続するアミノ酸配列からなるエピトープに結合するモノクローナル抗体は、断片が該連続するアミノ酸配列からなるエピトープを含んでいる場合には、不連続なアミノ酸配列からなるエピトープに結合するモノクローナル抗体よりも、該断片の1つの連続するアミノ酸配列に強く結合して該断片とIL-33受容体との結合を阻害等することから有利である。しかしながら、所望のアンタゴニスト作用を有するIL-33モノクローナル抗体を生成するための、このような連続するアミノ酸配列からなるエピトープを特定することは依然として困難であった。 【0008】 IL-33の連続するアミノ酸配列からなるエピトープに結合する抗IL-33中和モノクローナル抗体としては、ヒト等に投与した場合に抗原性が低いことが望ましい。ヒト抗体であれば、ヒトに投与した場合に抗原性が低く、さらにフレームワーク領域がヒト生殖系列のアミノ酸配列またはその組合せのアミノ酸配列であることが望ましい。しかしながら、ヒト抗体遺伝子ライブラリ中に含まれるヒト抗体にSHM-XELプラットフォーム等を適用した場合には、相補性決定領域に限らずフレームワーク領域にもアミノ酸配列の変異が入ってしまう。また、ヒト抗体遺伝子を導入したマウスをヒトIL-33タンパク質で免疫してヒト抗IL-33中和モノクローナル抗体を取得する場合にも、該抗IL-33中和モノクローナル抗体のフレームワーク領域にアミノ酸配列の変異が生じることを防止できない。したがって、フレームワーク領域のアミノ酸配列がヒト生殖系列のアミノ酸配列またはその組み合わせのアミノ酸配列である、IL-33に対する単離したヒトモノクローナル抗体の取得は依然として困難であった。 【課題を解決するための手段】 【0009】 本発明者らは、上記課題を解決すべく、鋭意検討した結果、従来の好ましいエピトープであると考えられていた155~198位に存在するエピトープに強く結合する抗体が、ほとんどアンタゴニスト作用を有さない一方で、101位~154位又は199位~270位に存在する連続するアミノ酸配列からなるエピトープ、特に111~130位、131~150位、231~250位、又は251~270位に存在する連続するアミノ酸配列からなるエピトープが、該エピトープに対し結合する抗体のアンタゴニスト作用の観点で重要であることを見出し、本発明に至った。 【0010】 さらに本発明者らは、ヒト抗体ライブラリからヒト抗IL-33中和モノクローナル抗体を単離し、さらにその相補性決定領域にのみ変異を導入することにより結合性が高く、物性のよい相補性決定領域を特定した。これにより、生殖系列のフレームワーク領域のアミノ酸配列に変異を含まないヒト抗体であって、ヒトIL-33に結合し、その機能を中和するヒト抗体を取得できた。そこで本発明は以下の発明に関する: (【0011】以降は省略されています) この特許をJ-PlatPatで参照する
 特許ウォッチ
特許ウォッチ