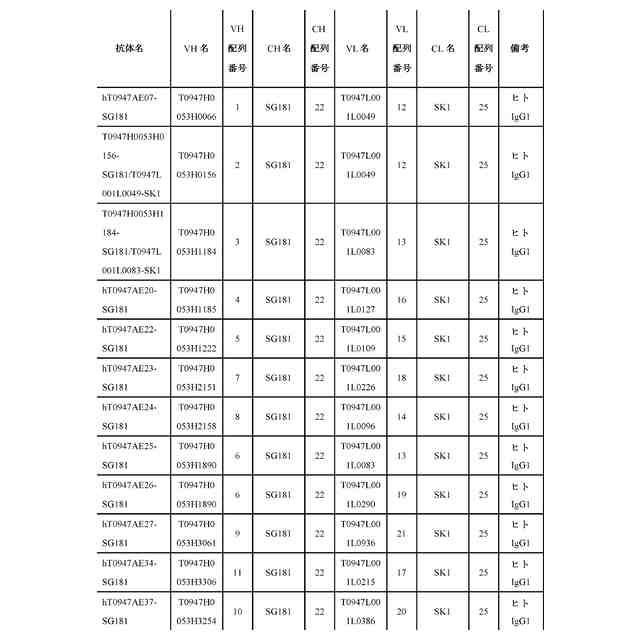
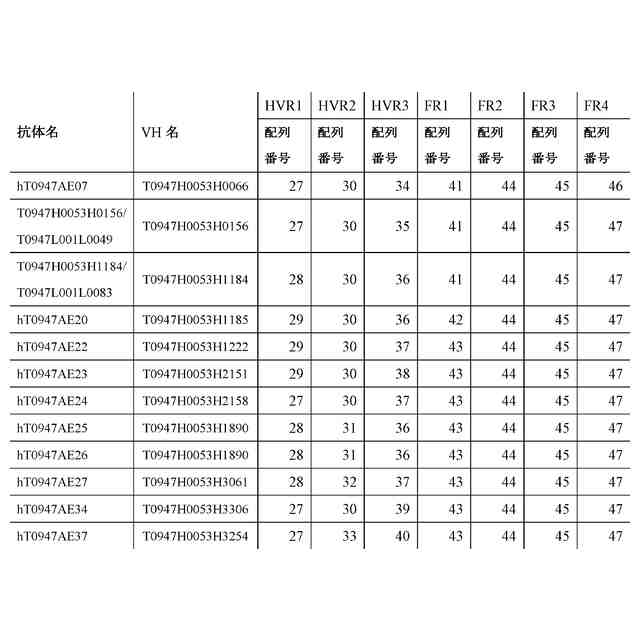
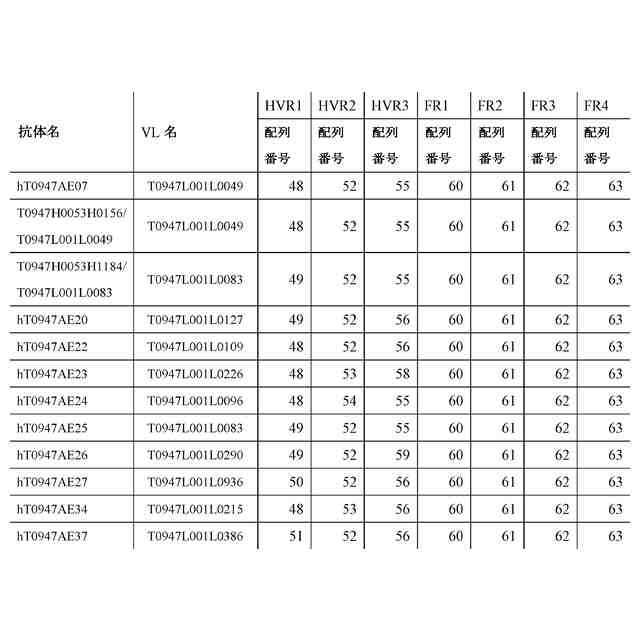
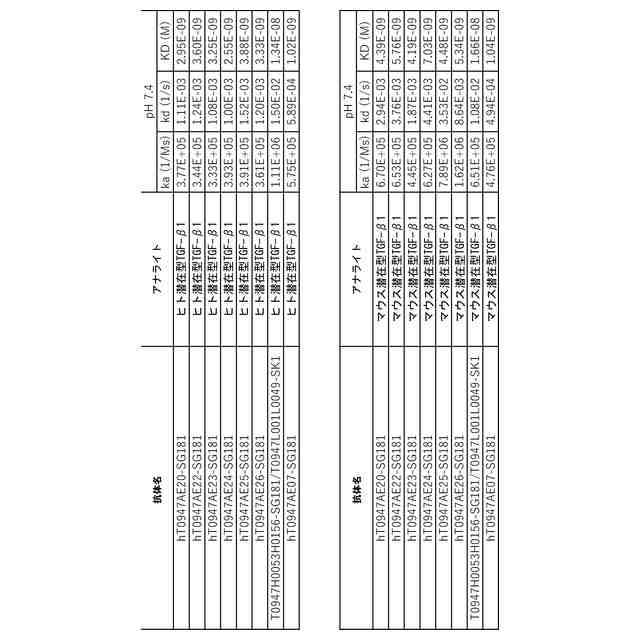
公開番号2024119756 公報種別公開特許公報(A) 公開日2024-09-03 出願番号2024019137 出願日2024-02-13 発明の名称抗潜在型TGF-β1抗体および使用方法 出願人中外製薬株式会社 代理人個人,個人,個人,個人,個人,個人,個人,個人,個人,個人,個人,個人 主分類C07K 16/22 20060101AFI20240827BHJP(有機化学) 要約【課題】本発明の目的は、抗潜在型TGF-β1抗体およびそのような抗体を用いる方法を提供することである。 【解決手段】本発明が提供する抗潜在型TGF-β1抗体は、潜在型TGF-β1のインテグリン媒介活性化を阻害することなくもしくは潜在型TGF-β1のインテグリン媒介活性化を部分的に阻害しつつ潜在型TGF-β1のプロテアーゼ媒介活性化を阻害するものであるか、または潜在型TGF-β1のプロテアーゼ媒介活性化を阻害するが、潜在型TGF-β1のインテグリン媒介活性化の阻害に対する影響がより小さいものである。本発明はまた、pH依存的結合特性が改善されている最適化された抗潜在型TGF-β1抗体も提供する。抗潜在型TGF-β1抗体は、線維症または癌を治療するために用いられてもよい。例えば、腎線維症、肝線維症、および肺線維症が、抗潜在型TGF-β1抗体を用いて治療されてもよい。 【選択図】なし 特許請求の範囲【請求項1】 以下を含む、抗潜在型TGF-β1抗体: (a)配列番号:29、30、および36のアミノ酸配列をそれぞれ含むHVR-H1、HVR-H2、およびHVR-H3、ならびに配列番号:49、52、および56のアミノ酸配列をそれぞれ含むHVR-L1、HVR-L2、およびHVR-L3; (b)配列番号:28、32、および37のアミノ酸配列をそれぞれ含むHVR-H1、HVR-H2、およびHVR-H3、ならびに配列番号:50、52、および56のアミノ酸配列をそれぞれ含むHVR-L1、HVR-L2、およびHVR-L3; (c)配列番号:27、30、および39のアミノ酸配列をそれぞれ含むHVR-H1、HVR-H2、およびHVR-H3、ならびに配列番号:48、53、および56のアミノ酸配列をそれぞれ含むHVR-L1、HVR-L2、およびHVR-L3;または (d)配列番号:27、33、および40のアミノ酸配列をそれぞれ含むHVR-H1、HVR-H2、およびHVR-H3、ならびに配列番号:51、52、および56のアミノ酸配列をそれぞれ含むHVR-L1、HVR-L2、およびHVR-L3。 続きを表示(約 1,400 文字)【請求項2】 以下を含む、請求項1記載の抗潜在型TGF-β1抗体: (a)(i)配列番号:4のアミノ酸配列と少なくとも95%の配列同一性を有するVH配列、(ii)配列番号:16のアミノ酸配列と少なくとも95%の配列同一性を有するVL配列、もしくは(iii)(i)におけるVH配列および(ii)におけるVL配列; (b)(i)配列番号:9のアミノ酸配列と少なくとも95%の配列同一性を有するVH配列、(ii)配列番号:21のアミノ酸配列と少なくとも95%の配列同一性を有するVL配列、もしくは(iii)(i)におけるVH配列および(ii)におけるVL配列; (c)(i)配列番号:11のアミノ酸配列と少なくとも95%の配列同一性を有するVH配列、(ii)配列番号:17のアミノ酸配列と少なくとも95%の配列同一性を有するVL配列、もしくは(iii)(i)におけるVH配列および(ii)におけるVL配列;または (d)(i)配列番号:10のアミノ酸配列と少なくとも95%の配列同一性を有するVH配列、(ii)配列番号:20のアミノ酸配列と少なくとも95%の配列同一性を有するVL配列、もしくは(iii)(i)におけるVH配列および(ii)におけるVL配列。 【請求項3】 以下を含む、請求項1記載の抗潜在型TGF-β1抗体: (a)配列番号:4のVH配列および配列番号:16のVL配列; (b)配列番号:9のVH配列および配列番号:21のVL配列; (c)配列番号:11のVH配列および配列番号:17のVL配列;または (d)配列番号:10のVH配列および配列番号:20のVL配列。 【請求項4】 ヒト抗体、ヒト化抗体、またはキメラ抗体である、請求項1記載の抗潜在型TGF-β1抗体。 【請求項5】 IgG抗体である、請求項1記載の抗潜在型TGF-β1抗体。 【請求項6】 野生型IgG1 Fc領域と比較して減少したエフェクター機能を有する改変IgG1 Fc領域を含む、請求項1記載の抗潜在型TGF-β1抗体。 【請求項7】 潜在型TGF-β1にpH依存的に結合する、請求項1記載の抗潜在型TGF-β1抗体。 【請求項8】 以下を含む、抗潜在型TGF-β1抗体: (a)配列番号:82の重鎖配列および配列番号:83の軽鎖配列; (b)配列番号:84の重鎖配列および配列番号:85の軽鎖配列; (c)配列番号:86の重鎖配列および配列番号:87の軽鎖配列; (d)配列番号:88の重鎖配列および配列番号:89の軽鎖配列; (e)配列番号:90の重鎖配列および配列番号:83の軽鎖配列; (f)配列番号:91の重鎖配列および配列番号:85の軽鎖配列; (g)配列番号:92の重鎖配列および配列番号:87の軽鎖配列;または (h)配列番号:93の重鎖配列および配列番号:89の軽鎖配列。 【請求項9】 請求項1記載の抗潜在型TGF-β1抗体と細胞傷害剤とを含む、イムノコンジュゲート。 【請求項10】 請求項1記載の抗潜在型TGF-β1抗体をコードする、単離された核酸。 (【請求項11】以降は省略されています) 発明の詳細な説明【技術分野】 【0001】 本発明は、抗潜在型TGF-β1抗体およびその使用方法に関する。 続きを表示(約 5,300 文字)【背景技術】 【0002】 形質転換成長因子β(形質転換成長因子ベータ;TGF-β)は、TGF-βアイソフォーム、アクチビン、インヒビン、Nodal、骨形成タンパク質(BMP)、抗ミュラー管ホルモン(AMH)および増殖分化因子(GDF)からなる、サイトカインのTGF-βスーパーファミリーのメンバーである。このスーパーファミリーのメンバーは、保存された構造を有する二量体タンパク質であり、インビトロおよびインビボで多面的な機能を有する(非特許文献1、2)。TGF-βアイソフォームは、増殖阻害、細胞遊走、浸潤、上皮間葉転換(EMT)、細胞外マトリクス(ECM)リモデリングおよび免疫抑制を含む多くの細胞プロセスに関与する(非特許文献3)。しかし、通常時には動的に調節され、組織の恒常性に関与しているにもかかわらず、TGF-βアイソフォームはしばしば、癌、線維症および炎症を含む疾患状態において慢性的に過剰発現され、このTGF-βの過剰産生は、細胞増殖、遊走または表現型を変化させることにより疾患の進行を促進する。 【0003】 哺乳動物においては3つの別個のTGF-βアイソフォーム(TGF-β1、TGF-β2およびTGF-β3)が同定されており、これらはアミノ酸レベルで70~82%の相同性を共有している(非特許文献4)。3つのTGF-βアイソフォームはすべて、ホモ二量体(それらの活性形態)でTGF-β受容体2型(TGFR2)に結合し;TGFR2はそれにより、TGF-β受容体1型(TGFR1)を動員および活性化して、受容体シグナル伝達を活性化させる(非特許文献5)。しかし、3つのアイソフォームの発現レベルは、組織によって様々であり(非特許文献6)、それらの機能は、ノックアウトマウスの表現型によって示されるように、異なっている(非特許文献7~11)。 【0004】 TGF-βスーパーファミリーの他のメンバーと同様、TGF-βは、その潜在関連ペプチド(latency-associated peptide; LAP)および潜在型TGF-β結合タンパク質(LTBP)と相互作用して潜在型大複合体(large latent complex; LLC)と呼ばれる大きな複合体を形成するホモ二量体を形成する前駆体タンパク質として合成される。TGF-β遺伝子は、シグナルペプチド、プロタンパク質コンバターゼ(PPC)切断部位で終了するプロペプチド、および成熟TGF-β配列からなるプレプロタンパク質配列をコードする。フーリンは、PPC切断部位を加水分解し、TGF-βおよびプロペプチド由来の別々のホモ二量体を生成する。これら2つのホモ二量体は、非共有結合的に結合した状態を維持し、分泌される。この潜在型複合体は、TGF-βを、その受容体に結合することができない不活性形態で維持する(非特許文献12、13)。TGF-β活性化プロセスは、ECMからのLLCの放出、その後のLAPのさらなるタンパク質分解によるその受容体への活性TGF-βの放出を含む(非特許文献3)。潜在型TGF-βは、プラスミン(PLN)、血漿カリクレイン(PLK)、マトリクスメタロプロテイナーゼ(MMP)2およびMMP9を含む広範囲のプロテアーゼ(非特許文献14)により、ならびにトロンボスポンジン1(TSP-1)(非特許文献15)により切断され活性TGF-βを放出する。何らかの学説に拘束されることを望まないが、MMP2およびMMP9は、潜在型TGF-β1をタンパク質分解により切断し、潜在型形態から成熟TGF-β1を放出する。MMP2およびMMP9は両方とも、不活性なプロMMPとして合成される。プロMMP2は、膜型MMP1(MT1-MMP/MMP14)および組織メタロプロテアーゼ阻害物質2(TIMP-2)の複合体によって活性化される。プロMMP9は、プラスミンおよびストロメリシン1(MMP-3)を含む相互作用プロテアーゼカスケードを通じて活性化される。プラスミンは、その酵素原から活性MMP-3を生成する。活性MMP-3は、92-kDaのプロMMP-9からプロペプチドを切断し、82-kDaの酵素的に活性な酵素を生じる。MMPの切断部位は、具体的に決定されていないが;MMP3は、潜在型TGF-βの79 Alaと80 Leuの間の部位を特異的に切断し、TGF-βを活性化することが報告されている(WO2005/023870)。あるいは、機械的伸展により、インテグリンが、LAP内に存在するRGDモチーフへの結合によりTGF-βを活性化し、その潜在型複合体からの成熟TGF-βの放出を誘導し得る(非特許文献16、17)。 【0005】 活性化後、二量体TGF-βリガンドは、I型およびII型受容体の細胞外ドメインに結合して近接するよう誘導し、受容体の細胞内セリン/スレオニンキナーゼドメインを、I型受容体のリン酸化およびその後の活性化を促進する立体配置にする。このI型受容体の活性化は、少なくとも2つの表面上独立している経路:SMAD依存的カノニカル経路およびSMAD非依存的または非カノニカル経路によるシグナル伝達の増幅をもたらす。SMAD依存的経路において、TGFR1(ALK5としても公知)の活性化は、SMADタンパク質のリン酸化をもたらす。SMAD2およびSMAD3は、TGFR1の基質である。受容体によりリン酸化されると、SMADは共通のメディエーターであるSMAD4とともに核に移行し、そこでそれらは他の転写因子と相互作用して転写応答を調節する(非特許文献18)。非カノニカル経路において、活性化されたTGF-β受容体複合体は、他の因子、例えば腫瘍壊死因子(TNF)受容体関連因子4(TRAF4)、TRAF6、TGF-β活性化キナーゼ1(TAK1、MAP3K7としても公知)、p38マイトジェン活性化タンパク質キナーゼ(p38 MAPK)、RHO、ホスホイノシチド3-キナーゼ(PI3K)、AKT(プロテインキナーゼBとしても公知)、細胞外シグナル調節キナーゼ(ERK)、JUN N末端キナーゼ(JNK)または核因子κB(NF-κB)を通じてシグナルを伝達する。したがって、TGF-βシグナル伝達に対する細胞応答は、カノニカルおよび非カノニカルシグナル伝達カスケードの動的な連携によりもたらされる。 【0006】 線維症、または瘢痕組織を形成するECM分子の蓄積は、慢性組織損傷の一般的な特徴である。腎線維症、肝線維症、および肺(lung/pulmonary)線維症は、特により一般的な線維症であり、これらに対してアンメットメディカルニーズが多く存在する。TGF-βは、間葉系細胞の細胞外マトリクスの生成を強く促進しつつ、同時に上皮細胞の増殖を抑制し、これが硬化性疾患の発病に寄与している。トランスジェニックマウスの肝臓における活性形態のTGF-β1の過剰発現は、複数の臓器において線維症を誘導するのに十分である(非特許文献19)。他方、TGF-βはまた、我々の健康の維持においても重要な役割を担っている。例えば、TGF-βは、肺においてプロテアーゼの過剰生成を抑制し、肺気腫を引き起こす肺組織の破壊を抑制する。また、TGF-β1が欠如したマウスは、出生前致死(交尾後10.5日で約50%)または出生後、肺(血管炎、血管周囲性細胞浸潤、および間質性肺炎)ならびに心臓(心内膜炎および心筋炎)を含む多くの臓器において重度の炎症病巣が見られ、まもない死を示す。このことは、TGF-β1が免疫恒常性の維持において決定的な役割を担っていることを示唆している(非特許文献7)。 【0007】 TGF-βに対する中和抗体および動物モデルを用いた研究の結果は、TGF-βの作用を抑制することにより硬化性疾患を予防または治癒することができることを明らかにした。TGF-βは前駆体タンパク質として産生されることから、その潜在形態からの活性化を防止するアプローチがいくつか報告されている。その潜在形態からの活性化を防止する別の方法は、潜在型TGF-βに結合する阻害剤または抗体を使用して、プロテアーゼ、例えばPLKおよびPLNによる切断をブロックすることである。TGF-βの活性化を抑制するこの方法を使用する様々な抗体が、肝硬変を予防または治療することが報告された(特許文献1)。加えて、いくつかの文献において、癌を治療するための抗LAP抗体(特許文献2)およびTGF-β1関連障害を治療するためのTGF-β1結合免疫グロブリン(特許文献3)が言及されている。いくつかの抗潜在型TGF-β1抗体が、既に記述されている(特許文献4、特許文献5、および特許文献6)。 【先行技術文献】 【特許文献】 【0008】 WO 2011102483 WO 2016115345 WO 2017156500 WO 2018/043734 WO 2019/163927 WO 2021/039945 【非特許文献】 【0009】 McCartney-Francis, N. L. et al. Int. Rev. Immunol. 16, 553-580 (1998) Massague, J. Annu. Rev. Biochem. 67, 753-791 (1998) Derynck, R. & Miyazono, K. Cold Spring Harbor Press (2008) Yu, L. et al. Kidney Int. 64, 844-856 (2003). Xu, P., Liu, J. & Derynck, R. et al. FEBS Lett. 586, 1871-1884 (2012). Millan, F. A. et al. Development 111, 131-143 (1991). Kulkarni, A. B. et al. Proc. Natl Acad. Sci. USA 90, 770-774 (1993). Shull, M. M. et al. Nature 359, 693-699 (1992). Dickson, M. C. et al. Development 121, 1845-1854 (1995). Sanford, L. P. et al. Development 124, 2659-2670 (1997). Proetzel, G. et al. Nature Genet. 11, 409-414 (1995). Dubois, C. M.et al. J. Biol. Chem. 270, 10618-10624 (1995) Nunes, I. et al. J. Am. Optom. Assoc. 69, 643-648 (1998) Annes, J. et al. J. Cell Sci. 116, 217-224 (2003). Schultz-Cherry, S. et al. J. Biol. Chem. 269, 26775-26782 (1994). Munger, J. S. et al. Cell 96, 319-328 (1999). Shi, M. et al. Nature 474, 343-349 (2011). Shi, Y. & Massague, et al. Cell 113, 685-700 (2003). Sanderson, N. et al. Proc. Natl Acad. Sci.USA 92, 2572-2576 (1995). 【発明の概要】 【発明が解決しようとする課題】 【0010】 本発明の一つの目的は、潜在型TGF-β1のインテグリン媒介活性化を阻害することなくもしくは潜在型TGF-β1のインテグリン媒介活性化を部分的に阻害しつつ潜在型TGF-β1のプロテアーゼ媒介活性化を阻害するか、または潜在型TGF-β1のプロテアーゼ媒介活性化を阻害するが、潜在型TGF-β1のインテグリン媒介活性化の阻害に対する影響がより小さい、種交差性の(cross-species)ヒト化および最適化された抗潜在型TGF-β1抗体を提供することである。本発明はまた、潜在型TGF-β1に対してpH依存性結合アフィニティを有する抗TGF-β1抗体も提供する。 【課題を解決するための手段】 (【0011】以降は省略されています) この特許をJ-PlatPatで参照する





 特許ウォッチ
特許ウォッチ