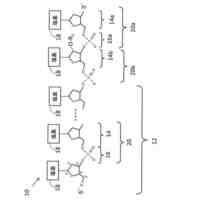
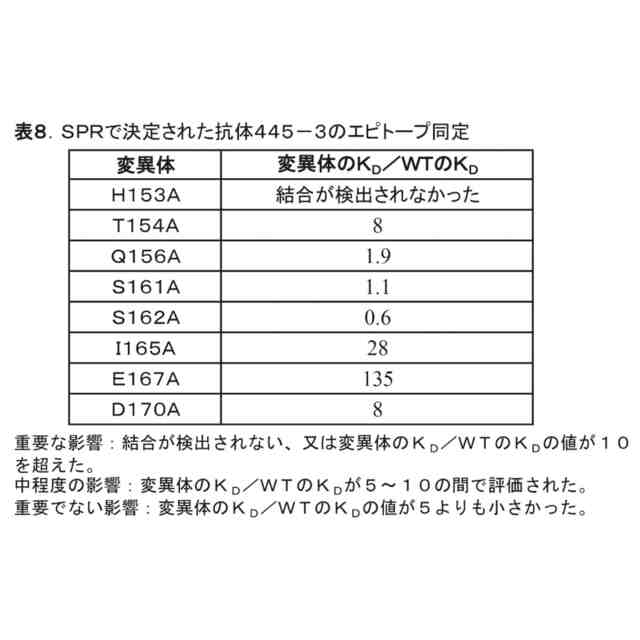
公開番号2024056938 公報種別公開特許公報(A) 公開日2024-04-23 出願番号2024022697,2020565492 出願日2024-02-19,2019-05-22 発明の名称抗OX40抗体及び使用方法 出願人ベイジーン リミテッド 代理人弁理士法人坂本国際特許商標事務所 主分類C12N 15/13 20060101AFI20240416BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学) 要約【課題】OX40のOX40Lとの相互作用を妨害せずに、OX40に特異的に結合するOX40アゴニスト抗体を提供すること。 【解決手段】ヒトOX40(ACT35、CD134又はTNFRSF4)に結合する抗体又はその抗原結合フラグメント、この抗体を含む医薬組成物、及び癌等の疾患を治療するためのこの抗体の使用又は組成物である。 【選択図】図1 特許請求の範囲【請求項1】 (i)(a)配列番号3のHCDR(重鎖相補性決定領域)1、(b)配列番号24のHCDR2、(c)配列番号5のHCDR3を含む重鎖可変領域、及び(d)配列番号25のLCDR(軽鎖相補性決定領域)1、(e)配列番号19のLCDR2、(f)配列番号8のLCDR3を含む軽鎖可変領域、 (ii)(a)配列番号3のHCDR1、(b)配列番号18のHCDR2、(c)配列番号5のHCDR3を含む重鎖可変領域、及び(d)配列番号6のLCDR1、(e)配列番号19のLCDR2、(f)配列番号8のLCDR3を含む軽鎖可変領域、 (iii)(a)配列番号3のHCDR1、(b)配列番号13のHCDR2、(c)配列番号5のHCDR3を含む重鎖可変領域、及び(d)配列番号6のLCDR1、(e)配列番号7のLCDR2、(f)配列番号8のLCDR3を含む軽鎖可変領域、又は (iv)(a)配列番号3のHCDR1、(b)配列番号4のHCDR2、(c)配列番号5のHCDR3を含む重鎖可変領域、及び(d)配列番号6のLCDR1、(e)配列番号7のLCDR2、(f)配列番号8のLCDR3を含む軽鎖可変領域を含み、 ヒトOX40に特異的に結合する抗体又はその抗原結合フラグメント。 続きを表示(約 1,700 文字)【請求項2】 (i)配列番号26と少なくとも90%、91%、92%、93%、94%、95%、96%、97%、98%若しくは99%同一のアミノ酸配列を含む重鎖可変領域(VH)、及び配列番号28と少なくとも90%、91%、92%、93%、94%、95%、96%、97%、98%若しくは99%同一のアミノ酸配列を含む軽鎖可変領域(VL)、 (ii)配列番号20と少なくとも90%、91%、92%、93%、94%、95%、96%、97%、98%若しくは99%同一のアミノ酸配列を含む重鎖可変領域(VH)、及び配列番号22と少なくとも90%、91%、92%、93%、94%、95%、96%、97%、98%若しくは99%同一のアミノ酸配列を含む軽鎖可変領域(VL)、 (iii)配列番号14と少なくとも90%、91%、92%、93%、94%、95%、96%、97%、98%若しくは99%同一のアミノ酸配列を含む重鎖可変領域(VH)、及び配列番号16と少なくとも90%、91%、92%、93%、94%、95%、96%、97%、98%若しくは99%同一のアミノ酸配列を含む軽鎖可変領域(VL)、又は (iv)配列番号9と少なくとも91%、92%、93%、94%、95%、96%、97%、98%若しくは99%同一のアミノ酸配列を含む重鎖可変領域(VH)、及び配列番号11と少なくとも91%、92%、93%、94%、95%、96%、97%、98%若しくは99%同一のアミノ酸配列を含む軽鎖可変領域(VL)を含む、 請求項1に記載の抗体又はその抗原結合フラグメント。 【請求項3】 配列番号26、28、20、22、14、16、9及び/又は11の中の1、2、3、4、5、6、7、8、9又は10個のアミノ酸が、挿入、削除又は置換されている、請求項2に記載の抗体又はその抗原結合フラグメント。 【請求項4】 (i)配列番号26を含む重鎖可変領域(VH)、及び配列番号28を含む軽鎖可変領域(VL)、 (ii)配列番号20を含む重鎖可変領域(VH)、及び配列番号22を含む軽鎖可変領域(VL)、 (iii)配列番号14を含む重鎖可変領域(VH)、及び配列番号16を含む軽鎖可変領域(VL)、又は (iv)配列番号9を含む重鎖可変領域(VH)、及び配列番号11を含む軽鎖可変領域(VL)を含む、 請求項1に記載の抗体又はその抗原結合フラグメント。 【請求項5】 モノクローナル抗体、キメラ抗体、ヒト化抗体、ヒト遺伝子操作抗体、単鎖抗体(scFv)、Fabフラグメント、Fab’フラグメント、又はF(ab’) 2 フラグメントである、請求項1~4のいずれかに記載の抗体又はその抗原結合フラグメント。 【請求項6】 OX40アゴニスト活性を有する、請求項1~5のいずれかに記載の抗体又はその抗原結合フラグメント。 【請求項7】 ヒトOX40のH153~D170からなる群から選択される1以上のアミノ酸残基を含むエピトープでヒトOX40に結合する、請求項1~6のいずれかに記載の抗体又はその抗原結合フラグメント。 【請求項8】 ヒトOX40のH153、T154、I165、E167及びD170からなる群から選択される1以上のアミノ酸残基を含むエピトープでヒトOX40に結合する、請求項1~6のいずれかに記載の抗体又はその抗原結合フラグメント。 【請求項9】 ヒトOX40のH153、I165及びE167からなる群から選択される1以上のアミノ酸残基を含むエピトープでヒトOX40に結合する、請求項1~6のいずれかに記載の抗体又はその抗原結合フラグメント。 【請求項10】 配列番号30で、又は配列番号30の内部でヒトOX40に結合する、請求項1~6のいずれかに記載の抗体又はその抗原結合フラグメント。 (【請求項11】以降は省略されています) 発明の詳細な説明【技術分野】 【0001】 本明細書には、ヒトOX40に結合する抗体又はその抗原結合フラグメント、この抗体を含む組成物、及び癌の治療に使用する方法が開示される。 続きを表示(約 4,300 文字)【背景技術】 【0002】 OX40(ACT35、CD134又はTNFRSF4とも呼ばれる)は、約50KDのI型膜貫通糖タンパク質であり、腫瘍壊死因子受容体スーパーファミリー(TNFRSF)の一員である(Croft, 2010;Gough and Weinberg, 2009)。成熟したヒトOX40は、249アミノ酸(AA)残基で構成され、37AAの細胞質側末端と185AAの細胞外領域を有する。OX40の細胞外ドメインは、3つの完全なシステインリッチドメイン(CRD)と1つの不完全なCRDを含む。OX40の細胞内ドメインは、1つの保存されたシグナル伝達関連QEEモチーフを含み、これは、TRAF2、TRAF3及びTRAF5を含むいくつかのTNFR関連因子(TRAF)への結合を媒介し、OX40が細胞内キナーゼに結合できるようにする(Arch and Thompson, 1998;Willoughby et al., 2017)。 【0003】 OX40は、活性化されたラットCD4 + T細胞上で最初に発見され、その後、T細胞からネズミとヒトの相同体がクローン化された(al-Shamkhani et al., 1996;Calderhead et al., 1993)。Tヘルパー(Th)1細胞、Th2細胞、Th17細胞及び制御性T(Treg)細胞を含む、活性化されたCD4 + T細胞の上での発現に加えて、CD8 + T細胞、ナチュラルキラー(NK)T細胞、好中球及びNK細胞の表面上でも、OX40発現が見られる(Croft, 2010)。対照的に、ナイーブCD4 + T細胞、ナイーブCD8 + T細胞、及びほとんどの休止メモリーT細胞の上で、低いOX40発現が見られる(Croft, 2010;Soroosh et al., 2007)。ナイーブT細胞上でのOX40の表面発現は一過性である。TCRの活性化後、T細胞上のOX40の発現は、24時間以内に大幅に増加し、2~3日でピークに達し、5~6日間持続する(Gramaglia et al., 1998)。 【0004】 OX40のリガンド(OX40L、gp34、CD252又はTNFSF4とも呼ばれる)は、OX40の唯一のリガンドである。TNFSF(腫瘍壊死因子スーパーファミリー)の他のメンバーと同様に、OX40LはII型糖タンパク質であり、183AAを含み、23AAの細胞内ドメイン及び133AAの細胞外ドメインを有する(Croft, 2010;Gough and Weinberg, 2009)。OX40Lは、細胞表面上にホモマー三量体複合体を自然に形成する。リガンド三量体は、主に受容体のCRD1、CRD2及び部分的なCRD3領域を介して、しかし、CRD4の関与無しに、リガンドモノマー-モノマーの境界面でOX40の3つの複製物と相互作用する(Compaan and Hymowitz, 2006)。OX40Lは、活性化された抗原提示細胞(APC)上で主に発現する。このAPCには、活性化されたB細胞(Stuber et al., 1995)、成熟した従来の樹状細胞(DC)(Ohshima et al., 1997)、形質細胞様樹状細胞(pDC)(Ito et al., 2004)、マクロファージ(Weinberg et al., 1999)及びランゲルハンス細胞(Sato et al., 2002)が含まれる。更に、OX40Lは、NK細胞、肥満細胞、活性化T細胞のサブセット、血管内皮細胞及び平滑筋細胞等の他の細胞種の上でも発現することが分かっている(Croft, 2010;Croft et al., 2009)。 【0005】 三量体のOX40Lによるライゲーション又はアゴニスト抗体による二量体化を経由するOX40の三量体化は、アダプター分子のTRAF2、TRAF3及び/又はTRAF5のリクルート及び細胞内QEEモチーフへのドッキングに寄与する(Arch and Thompson, 1998;Willoughby et al., 2017)。TRAF2及びTRAF3のリクルートとドッキングは、更に古典的NF-κB1経路及び非古典的NF-κB2経路の両方の活性化を導くことができる。これらの経路は、T細胞の生存、分化、増殖、サイトカイン産生及びエフェクター機能の調節に重要な役割を果たす(Croft, 2010;Gramaglia et al., 1998;Huddleston et al., 2006;Rogers et al., 2001;Ruby and Weinberg, 2009;Song et al., 2005a;Song et al., 2005b;Song et al., 2008)。 【0006】 OX40は正常組織では発現が低く、リンパ器官のリンパ球上で主に発現する(Durkop et al., 1995)。しかし、免疫細胞上でのOX40発現のアップレギュレーションが、動物モデルと、自己免疫疾患(Carboni et al., 2003;Jacquemin et al., 2015;Szypowska et al., 2014)及び癌(Kjaergaard et al., 2000;Vetto et al., 1997;Weinberg et al., 2000)等の病的状態(Redmond and Weinberg, 2007)のヒト患者の両方で頻繁に観察されている。特に、OX40の発現増加は、結腸直腸癌及び皮膚黒色腫の患者の生存期間の延長に関連しており、遠隔転移及びより進行した腫瘍の特徴の発生と逆相関する(Ladanyi et al., 2004;Petty et al., 2002;Sarff et al., 2008)。抗OX40抗体治療が、様々なマウスモデルで抗腫瘍効果を引き出すことができることも示され(Aspeslagh et al., 2016)、免疫療法のターゲットとしてのOX40の可能性が示された。Curti et alによって実施された最初の癌患者の臨床試験において、抗腫瘍効果及び腫瘍特異的T細胞の活性化の証拠が、アゴニスト性の抗OX40モノクローナル抗体を用いて観察され、OX40抗体が抗腫瘍T細胞応答を高める有用性を有することが示された(Curti et al., 2013)。 【0007】 抗腫瘍効果を媒介する際のアゴニスト性の抗OX40抗体の作用機序は、主にマウス腫瘍モデルで研究された(Weinberg et al., 2000)。最近まで、腫瘍におけるアゴニスト性の抗OX40抗体の作用機序は、エフェクターT細胞における共刺激シグナル伝達経路を誘発する能力、及びTreg細胞の分化と機能への抑制効果に帰された(Aspeslagh et al., 2016;Ito et al., 2006;St Rose et al., 2013;Voo et al., 2013)。最近の研究で、動物腫瘍モデルと癌患者の両方で、腫瘍浸潤性TregがエフェクターT細胞(CD4 + とCD8 + の両方)及び末梢Tregよりも高いレベルのOX40を発現することが示された(Lai et al., 2016;Marabelle et al., 2013b;Montler et al., 2016;Soroosh et al., 2007;Timperi et al., 2016)。従って、抗OX40抗体が抗腫瘍応答を誘発する二次的効果は、抗体依存性細胞傷害(ADCC)及び/又は抗体依存性細胞傷害(ADCC)を介して腫瘍内OX40 + Treg細胞を枯渇させる際のFc媒介エフェクター機能に依存する(Aspeslagh et al., 2016;Bulliard et al., 2014;Marabelle et al., 2013a;Marabelle et al., 2013b;Smyth et al., 2014)。この研究によって、Fc媒介エフェクター機能を有するアゴニスト性の抗OX40抗体が、腫瘍内Tregを優先的に枯渇させ、腫瘍微小環境におけるTregに対するCD8 + エフェクターT細胞の比率を改善することができ、その結果、抗腫瘍免疫応答が改善し、腫瘍の退縮が増加し、生存率が改善されることが示される(Bulliard et al., 2014;Carboni et al., 2003;Jacquemin et al., 2015;Marabelle et al., 2013b)。これらの発見に基づいて、アゴニスト性の活性及びFc媒介エフェクター機能の両方を備えるアゴニスト性の抗OX40抗体を開発するアンメット・メディカル・ニーズがある。 【発明の概要】 【発明が解決しようとする課題】 【0008】 今まで、病院で用いられるアゴニスト性の抗OX40抗体はほとんど、OX40-OX40Lの相互作用を遮断するリガンド競合性の抗体である(例えば、WO2016/196228A1)。OX40-OX40Lの相互作用は効果的な抗腫瘍免疫を高めるために不可欠であるため、OX40-OX40Lの遮断によって、これらのリガンド競合性の抗体の有効性が制限される。従って、OX40のOX40Lとの相互作用を妨害せずに、OX40に特異的に結合するOX40アゴニスト抗体は、癌及び自己免疫疾患の治療において有用性を有する。 【課題を解決するための手段】 【0009】 本開示は、OX40を活性化し、免疫細胞中のシグナル伝達を誘導し、従って抗腫瘍免疫を促進する、アゴニスト性の抗OX40抗体及びその抗原結合フラグメントに向けられる。 【0010】 1つの実施態様において、本開示は、ヒトOX40に結合するモノクローナル抗体又はその抗原結合フラグメントを提供する。1つの側面において、本開示の抗体は、OX40Lと競合しないか、又はOX40のそのリガンドOX40Lへの結合を妨害しない。 (【0011】以降は省略されています) この特許をJ-PlatPatで参照する









 特許ウォッチ
特許ウォッチ