TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025010204
公報種別
公開特許公報(A)
公開日
2025-01-20
出願番号
2024187836,2022111649
出願日
2024-10-25,2013-11-05
発明の名称
遺伝学的に修飾された非ヒト動物およびその使用法
出願人
リジェネロン・ファーマシューティカルズ・インコーポレイテッド
,
REGENERON PHARMACEUTICALS, INC.
,
イエール ユニバーシティ
,
インスティテュート フォー リサーチ イン バイオメディシン (アイアールビー)
代理人
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
主分類
A01K
67/0275 20240101AFI20250109BHJP(農業;林業;畜産;狩猟;捕獲;漁業)
要約
【課題】ヒト造血細胞の発達、機能、又は疾患をモデル化するために使用され得る、遺伝学的に修飾された非ヒト動物の提供。
【解決手段】IL-6プロモーターに機能的に連結されたヒトIL-6をコードする核酸を含む、遺伝学的に修飾された非ヒト動物。いくつかの場合において、ヒトIL-6を発現する遺伝学的に修飾された非ヒト動物は、ヒトM-CSF、ヒトIL-3、ヒトGM-CSF、ヒトSIRPa、又はヒトTPOのうちの少なくとも1種も発現する。
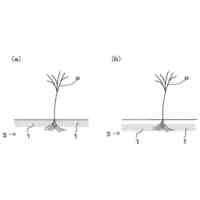
【選択図】図1
特許請求の範囲
【請求項1】
IL-6プロモーターに機能的に連結されたヒトIL-6をコードする核酸、ならびに
(i)非ヒト動物のゲノムにランダムに組み込まれた、SIRPaプロモーターに機能的に連結されたヒトSIRPaをコードする核酸;
(ii)M-CSFプロモーターに機能的に連結されたヒトM-CSFをコードする核酸;
(iii)IL-3プロモーターに機能的に連結されたヒトIL-3をコードする核酸;
(iv)GM-CSFプロモーターに機能的に連結されたヒトGM-CSFをコードする核酸;および
(v)TPOプロモーターに機能的に連結されたヒトTPOをコードする核酸:
からなる群より選択される少なくとも1種の付加的な核酸
を含む、遺伝学的に修飾された非ヒト動物。
続きを表示(約 730 文字)
【請求項2】
IL-6プロモーターが非ヒト動物IL-6プロモーターであり、ヒトIL-6をコードする核酸が非ヒト動物IL-6遺伝子座において非ヒト動物IL-6プロモーターに機能的に連結されている、請求項1に記載の遺伝学的に修飾された非ヒト動物。
【請求項3】
ヒトM-CSFをコードする核酸が非ヒト動物M-CSF遺伝子座にあり、ヒトIL-3をコードする核酸が非ヒト動物IL-3遺伝子座にあり、ヒトGM-CSFをコードする核酸が非ヒト動物GM-CSF遺伝子座にあり、ヒトTPOをコードする核酸が非ヒト動物TPO遺伝子座にある、請求項1に記載の遺伝学的に修飾された非ヒト動物。
【請求項4】
免疫不全である、請求項1に記載の遺伝学的に修飾された非ヒト動物。
【請求項5】
組換え活性化遺伝子(RAG)を発現しない、請求項4に記載の遺伝学的に修飾された非ヒト動物。
【請求項6】
IL2受容体γ鎖を発現しない(γ鎖-/-)、請求項4に記載の遺伝学的に修飾された非ヒト動物。
【請求項7】
RAG2を発現せず、かつIL2rgを発現しない、請求項4に記載の遺伝学的に修飾された非ヒト動物。
【請求項8】
ヒト造血細胞の生着をさらに含む、請求項1に記載の遺伝学的に修飾された非ヒト動物。
【請求項9】
ヒト造血細胞がCD34+細胞である、請求項8に記載の遺伝学的に修飾された非ヒト動物。
【請求項10】
ヒト造血細胞が多発性骨髄腫細胞である、請求項8に記載の遺伝学的に修飾された非ヒト動物。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
連邦政府によって後援された研究または開発に関する記述
本発明は、National Institutes of Health(NIH)によって授与されたグラント番号5R01CA156689-04の下で、政府の支援を受けてなされた。政府は、本発明における一定の権利を有する。
続きを表示(約 4,700 文字)
【0002】
関連出願の相互参照
本願は、35U.S.C.§119(e)に準じて、2012年11月5日出願の米国仮特許出願第61/722,437号の出願日に基づく優先権を主張するものであり、その全開示が参照によって本明細書に組み入れられる。
【背景技術】
【0003】
発明の背景
生物医学的研究の目標は、ヒト生理学についてのよりよい理解を獲得し、ヒト疾患を防止するか、処置するか、または治癒させるために、この知識を使用することである。ヒト対象における実験法に対する実務上および倫理上の障壁のため、多くの研究が、マウスのような小動物モデルにおいて実施されている。従って、これらのヒト疾患の動物モデルが必要とされている。
【0004】
例えば、米国内で、毎年およそ20,000人の患者が、抗体分泌高分化B細胞のほぼ不治の悪性疾患である多発性骨髄腫(MM)を有すると新たに診断される(Hideshima et al.,2007,Nat Rev Cancer.7:585-98(非特許文献1);Kuehl and Bergsagel,2002,Nat Rev Cancer.2:175-87(非特許文献2))。MMは、悪性形質細胞の骨髄(BM)への浸潤を特徴とし、臨床徴候には、骨疾患、高カルシウム血症、血球減少、腎機能障害、および末梢神経障害が含まれる(Hideshima et al.,2007,Nat Rev Cancer.7:585-98(非特許文献1);Kuehl and Bergsagel,2002,Nat Rev Cancer.2:175-87(非特許文献2))。50歳を超える者のおよそ3%に影響を与える良性単クローン性γグロブリン血症(MGUS)と呼ばれる前癌状態が、大部分の症例において、MMに先行する(Landgren et al.,2009,Blood 113:5412-7(非特許文献3))。核型の変化およびIgH転座を含む、複雑な不均一な遺伝学的異常が、MM細胞の特徴である(Kuehl and Bergsagel,2002,Nat Rev Cancer.2:175-87(非特許文献2);Zhan et al.,2006,Blood 108:2020-8(非特許文献4))。MGUSにおいて増幅される形質細胞クローンは、骨髄腫形質細胞に類似している遺伝子および表現型のプロファイルを有すると考えられる(Chng et al.,2005,Blood 106:2156-61(非特許文献5);Fonseca et al.,2002,Blood 100:1417-24(非特許文献6);Kaufmann et al.,2004,Leukemia.18:1879-82(非特許文献7))。サイクリンD遺伝子における変異が、MMを発症させることが示唆されているが、他の因子の可能性のある寄与は、決定的には証明されていない(Bergsagel et al.,2005,Blood 106:296-303(非特許文献8))。にもかかわらず、遺伝性の遺伝子改変は、MM細胞の挙動の唯一の決定因子ではない。むしろ、薬物に対する耐性およびサイトカインに対する異常な生物学的応答は、微小環境との相互作用によって強く影響を受けるため、それが、新規治療薬を開発する機会を与える。
【0005】
他の多くの腫瘍と同様に、MMは、支持的な環境を生み出す非悪性間質細胞と強く相互作用する不均一な細胞集団を特徴とする(De Raeve and Vanderkerken,2005,Histol Histopathol.20:1227-50(非特許文献9);Dhodapkar,2009,Am J Hematol.84:395-6(非特許文献10))。MM細胞のためのBM微小環境は、多様な細胞外マトリックス(ECM)、ならびに造血系起源および非造血系起源の両方の細胞成分からなる。BMは、正常な造血のための防御された環境を提供するが、ECMタンパク質および補助細胞とのMM細胞の相互作用が、MM病原において重大な役割を果たす(De Raeve and Vanderkerken,2005,Histol Histopathol.20:1227-50(非特許文献9);Dhodapkar,2009,Am J Hematol.84:395-6(非特許文献10);Hideshima et al.,2007,Nat Rev Cancer.7:585-98(非特許文献1))。間質細胞、骨髄系細胞、破骨細胞、および骨芽細胞は、MM細胞の遊走、生存、および増殖を媒介するシグナル経路を活性化する、インターロイキン6(IL-6)、B細胞活性化因子(BAFF)、繊維芽細胞増殖因子、および間質細胞由来因子1aのような増殖因子を産生する。特に、間質細胞、破骨細胞、および骨髄系細胞によって産生されるIL-6は、初期段階において、MMの病原のための重大な因子であるようである(De Raeve and Vanderkerken,2005,Histol Histopathol.20:1227-50(非特許文献9))。同様に、MM細胞との相互作用によって、破骨細胞および樹状細胞は、抗アポトーシスシグナルを提供するBAFFおよび/または増殖誘導リガンド(APRIL)を産生し、それらも薬物耐性を増加させる(De Raeve and Vanderkerken,2005,Histol Histopathol.20:1227-50(非特許文献9);Kukreja et al.,2006,J Exp Med.203:1859-65(非特許文献11))。
【0006】
癌の病原における主要なイベント(悪性細胞の調節されない増殖、生存、および蔓延)は、微小環境ニッチに存在する支持的な細胞型および可溶性因子の特定の組み合わせに依る。マウスモデルは、ヒトにおける悪性形質転換および疾患の推進力の重大な局面の特徴決定において重要な役割を果たす。しかしながら、それらがヒト疾患の遺伝学的複雑性および臨床病理学的特徴を表すことは稀である。免疫低下マウスへのヒト腫瘍の異種移植は広範に利用されているが、信頼性のある生着は、典型的には、高悪性度の腫瘍または細胞株によってのみ実現可能であった。
【0007】
ヒト腫瘍細胞を増殖させるために現在利用可能な最良のモデルは、B細胞、T細胞、およびNK細胞を欠く重症免疫不全マウスである。MMのケースにおいて、これらのマウスへの初代骨髄腫細胞の生着は成功していないが、初代骨髄腫細胞は、免疫低下マウスへの同時移植によって、ヒト胎児骨片に生着することができる(Yaccoby et al.,1998,Blood 92:2908-13(非特許文献12))。このモデルにおいて、MM細胞は、ヒト骨には見出されるが、マウス骨または末梢には検出されず、このことは、高度の残余の異種拒絶(xenorejection)およびヒトBM微小環境の必要性を証明している(Yaccoby et al.,1998,Blood 92:2908-13(非特許文献12);Yaccoby and Epstein,1999,Blood 94:3576-82(非特許文献13))。NOD/Scid/γc
-/-
マウスが数種のMM細胞株の生着を可能にすることが最近証明され、MMのインビボモデルとしての可能性が判明した(Dewan et al.,2004,Cancer Sci.95:564-8(非特許文献14);Miyakawa et al.,2004,Biochem Biophys Res Commun.313:258-62(非特許文献15))。しかしながら、低い異種拒絶を有するそれらのマウスモデルですら、種の壁を超えないが、形質転換細胞の増殖および生存を支持するために不可欠である多数の因子のため、制限された増殖環境を有する(Manz,2007)。腫瘍とその環境との間の複雑な病原性インタープレイを探究することを可能にするインビボモデルは、新たな薬物および治療を設計するために不可欠であろう。
【0008】
従って、初代ヒト造血器腫瘍細胞を含むヒト造血細胞をマウスにおいて確実に増殖させ研究するためのヒト化非ヒト動物および方法を開発する必要性は、未だ満たされていない。本発明は、当技術分野におけるこれらの満たされていない必要性を解決する。
【先行技術文献】
【非特許文献】
【0009】
Hideshima et al.,2007,Nat Rev Cancer.7:585-98
Kuehl and Bergsagel,2002,Nat Rev Cancer.2:175-87
Landgren et al.,2009,Blood 113:5412-7
Zhan et al.,2006,Blood 108:2020-8
Chng et al.,2005,Blood 106:2156-61
Fonseca et al.,2002,Blood 100:1417-24
Kaufmann et al.,2004,Leukemia.18:1879-82
Bergsagel et al.,2005,Blood 106:296-303
De Raeve and Vanderkerken,2005,Histol Histopathol.20:1227-50
Dhodapkar,2009,Am J Hematol.84:395-6
Kukreja et al.,2006,J Exp Med.203:1859-65
Yaccoby et al.,1998,Blood 92:2908-13
Yaccoby and Epstein,1999,Blood 94:3576-82
Dewan et al.,2004,Cancer Sci.95:564-8
Miyakawa et al.,2004,Biochem Biophys Res Commun.313:258-62
【発明の概要】
【0010】
ヒト造血細胞の発達、機能、または疾患をモデル化するために使用され得る、遺伝学的に修飾された非ヒト動物が提供される。遺伝学的に修飾された非ヒト動物は、IL-6プロモーターに機能的に連結されたヒトIL-6をコードする核酸を含む。いくつかの場合において、ヒトIL-6を発現する遺伝学的に修飾された非ヒト動物は、ヒトM-CSF、ヒトIL-3、ヒトGM-CSF、ヒトSIRPa、またはヒトTPOのうちの少なくとも1種も発現する。いくつかの場合において、遺伝学的に修飾された非ヒト動物は、免疫不全である。いくつかのそのような場合において、遺伝学的に修飾された非ヒト動物は、健常なまたは疾患を有するヒト造血細胞を生着させられる。ヒト造血細胞の発達、機能、および/または疾患のモデル化において本発明の遺伝学的に修飾された非ヒト動物を使用する方法、ならびに本発明の遺伝学的に修飾された非ヒト動物の作製および/または本発明の方法の実施において有用な試薬およびそのキットも、提供される。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
個人
除草具
2日前
個人
草抜き機
1か月前
個人
動物撃退装置
1か月前
個人
殺虫スプレー
13日前
個人
ルアー
27日前
個人
ベアスタンガン
2日前
個人
播種機と播種方法
28日前
井関農機株式会社
収穫機
5日前
井関農機株式会社
収穫機
5日前
個人
獣罠捕獲検知システム
2日前
井関農機株式会社
作業車両
13日前
個人
蔬菜栽培器
1か月前
個人
回転式草刈刃
1か月前
個人
根掛りしないイカ用疑似餌
2日前
個人
植物栽培装置
28日前
個人
植物支持装置
28日前
個人
動物の尿漏れ防止具
16日前
株式会社誠和
接触構造体
13日前
個人
有害獣類の忌避装置
7日前
株式会社ブリーデン
ルアー
27日前
日産化学株式会社
農薬粒状組成物
7日前
井関農機株式会社
作業車両
1か月前
井関農機株式会社
作業車両
13日前
株式会社天装
猫の爪研ぎ具
7日前
井関農機株式会社
作業車両
5日前
株式会社エスク
鳥獣忌避装置
2日前
株式会社サクマ
植物収容構造
5日前
有限会社トオヤ
魚釣り用ウキ
29日前
株式会社広瀬
植物の育成方法
1か月前
松山株式会社
農作業機
29日前
環境事業計画株式会社
動物用罠
7日前
東京都公立大学法人
摘み取り装置
1か月前
株式会社拓凌
動物捕獲用罠
7日前
井関農機株式会社
作業車両
5日前
株式会社ミラミラ
愛犬被服用型紙
1か月前
株式会社シマノ
魚釣用電動リール
1か月前
続きを見る
他の特許を見る










 特許ウォッチ
特許ウォッチ