TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
公開番号
2025009558
公報種別
公開特許公報(A)
公開日
2025-01-20
出願番号
2023112640
出願日
2023-07-07
発明の名称
アピイン生産のための新規遺伝子およびそれを用いたアピインの生産方法
出願人
学校法人立命館
,
学校法人常翔学園
代理人
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
主分類
C12N
9/10 20060101AFI20250110BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】本発明は、アピゲニンをアピインに変換するためのアピオース転移酵素をコードする遺伝子およびグルコース転移酵素をコードする遺伝子を同定し、当該遺伝子にコードされるアピオース転移酵素およびグルコース転移酵素を用いて、アピゲニンからアピインを一貫して生産するための方法を提供する。
【解決手段】特定のアミノ酸配列を含む、アピオース転移酵素を提供する。また、前記アピオース転移酵素、アピゲニン7-OグルコシドおよびUDP-アピオースを接触させる工程を含む、アピゲニン7-Oグルコシドからアピインを生産する方法を提供する。
【選択図】なし
特許請求の範囲
【請求項1】
以下の(i)~(iv)からなる群から選択されるアミノ酸配列を含む、アピオース転移酵素:
(i)配列番号2で表されるアミノ酸配列、
(ii)配列番号4で表されるアミノ酸配列、
(iii)配列番号2で表されるアミノ酸配列と実質的に同一のアミノ酸配列、
(iv)配列番号4で表されるアミノ酸配列と実質的に同一のアミノ酸配列。
続きを表示(約 1,200 文字)
【請求項2】
以下の(i)または(ii)のアミノ酸配列を含む、請求項1に記載のアピオース転移酵素:
(i)配列番号2で表されるアミノ酸配列、
(ii)配列番号4で表されるアミノ酸配列。
【請求項3】
以下の(i)~(iv)からなる群から選択されるアミノ酸配列を含む、グルコース転移酵素:
(i)配列番号6で表されるアミノ酸配列、
(ii)配列番号8で表されるアミノ酸配列、
(iii)配列番号6で表されるアミノ酸配列と実質的に同一のアミノ酸配列、
(iv)配列番号8で表されるアミノ酸配列と実質的に同一のアミノ酸配列。
【請求項4】
以下の(i)または(ii)のアミノ酸配列を含む、請求項3に記載のグルコース転移酵素:
(i)配列番号6で表されるアミノ酸配列、
(ii)配列番号8で表されるアミノ酸配列。
【請求項5】
請求項1または2に記載のアピオース転移酵素、アピゲニン7-OグルコシドおよびUDP-アピオースを接触させる工程を含む、アピゲニン7-Oグルコシドからアピインを生産する方法。
【請求項6】
該工程が、請求項1または2に記載のアピオース転移酵素をコードする塩基配列を有する核酸を含み、かつUDP-アピオースを生産する宿主細胞内で実施されることを特徴とする、請求項5に記載の方法。
【請求項7】
請求項3または4に記載のグルコース転移酵素、アピゲニンおよびUDP-グルコースを接触させる工程を含む、アピゲニンからアピゲニン7-Oグルコシドを生産する方法。
【請求項8】
該工程が、請求項3または4に記載のグルコース転移酵素をコードする塩基配列を有する核酸を含み、かつUDP-グルコースを生産する宿主細胞内で実施されることを特徴とする、請求項7に記載の方法。
【請求項9】
以下の(i)および(ii)の工程を含む、アピゲニンからアピインを生産する方法:
(i)請求項3または4に記載のグルコース転移酵素、アピゲニンおよびUDP-グルコースを接触させ、アピゲニン7-Oグルコシドを生産する工程、
(ii)請求項1または2に記載のアピオース転移酵素、該アピゲニン7-OグルコシドおよびUDP-アピオースを接触させ、アピインを生産する工程。
【請求項10】
工程(i)および(ii)が請求項3または4に記載のグルコース転移酵素をコードする塩基配列を有する核酸および請求項1または2に記載のアピオース転移酵素をコードする塩基配列を有する核酸を含み、かつUDP-グルコースおよびUDP-アピオースを生産する、同一の宿主細胞内で実施されることを特徴とする、請求項9に記載の方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、アピゲニンとUDP-グルコースからアピゲニン7-Oグルコシドを生産する活性を有する新規のグルコース転移酵素、アピゲニン7-OグルコシドとUDP-アピオースからアピインを生産する活性を有する新規のアピオース転移酵素、それらを用いたアピゲニンからアピインを生産する方法および生産キットに関する。
続きを表示(約 8,500 文字)
【背景技術】
【0002】
植物で生成されるフラボンの1種であるアピゲニンには、抗不安作用、がん細胞増殖抑制作用があり、糖尿病、健忘症、うつ病など有益な適応症が報告されている。これまでに大腸菌でのアピゲニン(非特許文献1)やアピゲニンの一つ手前の前駆体であるナリンゲニン(非特許文献2)の生産方法は報告があるものの、ヒトに有効に投与するにはアピゲニンに糖を結合させて可溶化させる必要があった。
【0003】
アピゲニンの配糖体の1種としてのアピインが知られており、セリ科のパセリやセロリに多く含まれることが知られている。アピインは、アピゲニンにUDP-グルコースをドナー基質として、グルコース転移酵素によって生合成されるアピゲニン7-Oグルコシドを経由し、さらにUDP-アピオースをドナー基質としてアピオース転移酵素によってアピゲニン7-Oグルコシドにアピオースが転移して生成されると考えられている。しかし、アピゲニン7-Oグルコシドからアピインの生成に至るためのアピオース転移酵素をコードする遺伝子は不明のままであるため、アピインの生産技術が確立されていなかった。
【先行技術文献】
【非特許文献】
【0004】
Miyahisa et al., Appl Microbiol Biotehcnol, 71, 53-58 (2006)
Zhou et al., J Microbiol Biotechnol, 30, 1574-1582 (2020)
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明の目的は、アピゲニンをアピインに変換するためのアピオース転移酵素をコードする遺伝子およびグルコース転移酵素をコードする遺伝子を同定し、当該遺伝子にコードされるアピオース転移酵素およびグルコース転移酵素を用いて、アピゲニンからアピインを一貫して生産するための方法を確立することである。
【課題を解決するための手段】
【0006】
発明者らは、パセリまたはセロリのRNA-Seq解析により、アピオース転移酵素候補遺伝子およびグルコース転移酵素候補遺伝子を選抜、クローニングし、その活性を確認することによって、新規のグルコース転移酵素をコードする遺伝子およびアピオース転移酵素をコードする遺伝子を同定することに成功した。また、発明者らは、同定したグルコース転移酵素をコードする遺伝子を大腸菌に導入して、得られた形質転換体から精製したグルコース転移酵素をアクセプター基質としてアピゲニンなどおよびドナー基質としてUDP-グルコースにin vitroで作用させることによって、アピゲニン7-O-グルコシドなどを生産させることに成功した。同様に、発明者らは、同定したアピオース転移酵素をコードする遺伝子を大腸菌に導入して、得られた形質転換体から精製したアピオース転移酵素をアクセプター基質としてアピゲニン7-O-グルコシドなどおよびドナー基質としてUDP-アピオースにin vitroで作用させることによって、アピインなどを生産させることに成功した。さらに、発明者らは、アピオース転移酵素をコードする遺伝子、グルコース転移酵素をコードする遺伝子を大腸菌に導入して、得られた形質転換体をアピゲニン存在下で培養することによって、アピゲニン7-O-グルコシドおよびアピインが生産されたことを確認した。
発明者らは、これらの知見に基づいてさらに検討を重ねた結果、本発明を完成するに至った。
【0007】
すなわち、本発明は、以下を提供する。
[1]以下の(i)~(iv)からなる群から選択されるアミノ酸配列を含む、アピオース転移酵素:
(i)配列番号2で表されるアミノ酸配列、
(ii)配列番号4で表されるアミノ酸配列、
(iii)配列番号2で表されるアミノ酸配列と実質的に同一のアミノ酸配列、
(iv)配列番号4で表されるアミノ酸配列と実質的に同一のアミノ酸配列。
[2]以下の(i)または(ii)のアミノ酸配列を含む、[1]に記載のアピオース転移酵素:
(i)配列番号2で表されるアミノ酸配列、
(ii)配列番号4で表されるアミノ酸配列。
[3]以下の(i)~(iv)からなる群から選択されるアミノ酸配列を含む、グルコース転移酵素:
(i)配列番号6で表されるアミノ酸配列、
(ii)配列番号8で表されるアミノ酸配列、
(iii)配列番号6で表されるアミノ酸配列と実質的に同一のアミノ酸配列、
(iv)配列番号8で表されるアミノ酸配列と実質的に同一のアミノ酸配列。
[4]以下の(i)または(ii)のアミノ酸配列を含む、[3]に記載のグルコース転移酵素:
(i)配列番号6で表されるアミノ酸配列、
(ii)配列番号8で表されるアミノ酸配列。
[5][1]または[2]に記載のアピオース転移酵素、アピゲニン7-OグルコシドおよびUDP-アピオースを接触させる工程を含む、アピゲニン7-Oグルコシドからアピインを生産する方法。
[6]該工程が、[1]または[2]に記載のアピオース転移酵素をコードする塩基配列を有する核酸を含み、かつUDP-アピオースを生産する宿主細胞内で実施されることを特徴とする、[5]に記載の方法。
[7][3]または[4]に記載のグルコース転移酵素、アピゲニンおよびUDP-グルコースを接触させる工程を含む、アピゲニンからアピゲニン7-Oグルコシドを生産する方法。
[8]該工程が、[3]または[4]に記載のグルコース転移酵素をコードする塩基配列を有する核酸を含み、かつUDP-グルコースを生産する宿主細胞内で実施されることを特徴とする、[7]に記載の方法。
[9]以下の(i)および(ii)の工程を含む、アピゲニンからアピインを生産する方法:
(i)[3]または[4]に記載のグルコース転移酵素、アピゲニンおよびUDP-グルコースを接触させ、アピゲニン7-Oグルコシドを生産する工程、
(ii)[1]または[2]に記載のアピオース転移酵素、該アピゲニン7-OグルコシドおよびUDP-アピオースを接触させ、アピインを生産する工程。
[10]工程(i)および(ii)が[3]または[4]に記載のグルコース転移酵素をコードする塩基配列を有する核酸および[1]または[2]に記載のアピオース転移酵素をコードする塩基配列を有する核酸を含み、かつUDP-グルコースおよびUDP-アピオースを生産する、同一の宿主細胞内で実施されることを特徴とする、[9]に記載の方法。
[11]以下の(i)~(iii)を含む、アピゲニン7-Oグルコシドからアピインの生産キット:
(i)[1]または[2]に記載のアピオース転移酵素、
(ii)アピゲニン7-Oグルコシド、
(iii)UDP-アピオース。
[12]以下の(i)および(ii)を含む、アピゲニン7-Oグルコシドからアピインの生産キット:
(i)[1]または[2]に記載のアピオース転移酵素をコードする塩基配列を有する核酸を含み、かつUDP-アピオースを生産する宿主細胞、
(ii)アピゲニン7-Oグルコシド。
[13]以下の(i)~(iii)を含む、アピゲニンからアピゲニン7-Oグルコシドの生産キット:
(i)[3]または[4]に記載のグルコース転移酵素、
(ii)アピゲニン、
(iii)UDP-グルコース。
[14]以下の(i)および(ii)を含む、アピゲニンからアピゲニン7-Oグルコシドの生産キット:
(i)[3]または[4]に記載のグルコース転移酵素をコードする塩基配列を有する核酸を含み、かつUDP-グルコースを生産する宿主細胞、
(ii)アピゲニン。
[15]以下の(i)~(v)を含む、アピゲニンからアピインの生産キット:
(i)[3]または[4]に記載のグルコース転移酵素、
(ii)[1]または[2]に記載のアピオース転移酵素、
(iii)アピゲニン、
(iv)UDP-グルコース、
(v)UDP-アピオース。
[16]以下の(i)および(ii)を含む、アピゲニンからアピインの生産キット:
(i)[3]または[4]に記載のグルコース転移酵素をコードする塩基配列を有する核酸および[1]または[2]に記載のアピオース転移酵素をコードする塩基配列を有する核酸を含み、かつUDP-グルコースおよびUDP-アピオースを生産する宿主細胞、
(ii)アピゲニン。
【発明の効果】
【0008】
本発明のアピオース転移酵素をコードする遺伝子およびグルコース転移酵素をコードする遺伝子を、UDP-グルコースおよびUDP-アピオースを生産する宿主細胞に導入し、アピゲニン存在下で培養することによって、アピゲニンからアピインを容易に生産することが可能となる。
【図面の簡単な説明】
【0009】
(A)AgApiTのアミノ酸配列を示す図である。C末端側の44アミノ酸はUGTに特徴的なPSPGモチーフである。(B)AgApiTの部分アミノ酸配列、セリ科のUGT94様タンパク質(UGT94P1、UGT89A2)の部分アミノ酸配列、Glehnia littoralis(ハマボウフウ)のGl16345の部分アミノ酸配列、Foeniculum vulgare(フェンネル)のFv4536、Fv22755.1およびFv18654.1の部分アミノ酸配列のアラインメントを示す図である。AgApiTの白抜きのIleは、ペントースの認識部位であると予想される。UGT94P1、UGT89A2の白抜きのIleは、ペントース認識部位のアミノ酸である。Gl16345、Fv4536、Fv22755.1およびFv18654.1の白抜きのValは、ペントース認識部位のアミノ酸である。
AgApiT発現コンストラクトを示す図である。AgApiTのN末端にHisタグ、ProS2タグがついている。
大腸菌で発現された組換えAgApiTのポリアクリルアミドゲル電気泳動像を示す図である。(-):発現誘導前の大腸菌培養液500 μLを遠心分離し、菌体を2×サンプルバッファー50 μLに溶かし、100 ℃で3分間インキュベート後、5 μLをアプライした。(+):0.8 mM IPTGを加え、15℃、24時間発現誘導後の大腸菌培養液を上記と同様に処理した。(AgApiT)計算分子量は49 kDa。(AgApiT+タグタンパク質)計算分子量は73 kDa。(タグタンパク質)計算分子量は24 kDa。
AgApiTの活性検出を示す図である。上段のクロマトグラムはネガティブコントロール1(タグタンパク質)、中段のクロマトグラムはネガティブコントロール2(ドナー基質UDP-Apiなし)、下段のクロマトグラムは2時間反応後の結果を示す。▼はアピイン、▽はアピゲニン7-O-β-D-グルコシドを示す。
AgApiTの酵素活性を示す図である。
(A)AgApiTによるアピイン生成反応の時間依存性を示す図である。(B)AgApiTによるアピイン生成反応の酵素量依存性を示す図である。
(A)アピゲニン7-O-β-D-グルコシドに対する反応速度を示す図である。(B)UDP-Apiに対する反応速度を示す図である。
AgApiTのドナー基質依存性を示す図である。50 μMアピゲニン7-O-β-D-グルコシド、0.5 mM糖ヌクレオチド、AgApiT 20 μgをpH 7.0、23℃で2時間反応させた。技術的反復実験を3回行い、結果の標準偏差をエラーバーとした。検出限界は20 pU/gである。
AgApiTのアクセプター基質依存性を示す図である。50 μMアクセプター基質、0.5 mM UDP-Api、AgApiT 20 μgをpH 7.0、23℃で2時間反応させた。技術的反復実験を3回行い、結果の標準偏差をエラーバーとした。検出限界は20 pU/gである。
AgApiTの様々なアクセプター基質に対する酵素活性を示す図である。50 μMアクセプター基質、0.5 mM UDP-Api、AgApiT 20 μgをpH 7.0、23℃で2時間反応させた。上段のクロマトグラムはネガティブコントロール(ドナー基質なし)、下段のクロマトグラムは2時間反応後の結果を示す。(A)アピゲニン7-O-β-D-グルコシド(B)ケルセチン7-O-β-D-グルコシド(C)ルテオリン7-O-β-D-グルコシド(D)ナリンゲニン7-O-β-D-グルコシド
AgApiTの点変異体のUDP-Apiに対する酵素活性を示す図である。50 μMアピゲニン7-O-β-D-グルコシド、0.5 mM UDP-Api、AgApiTの点変異体20 μgをpH 7.0、23°Cで2時間反応させた。技術的反復実験を3回行い、結果の標準偏差をエラーバーとした。検出限界は20 pU/gである。
セロリ発達段階ごとのAgApiT遺伝子の発現量を示す図である。
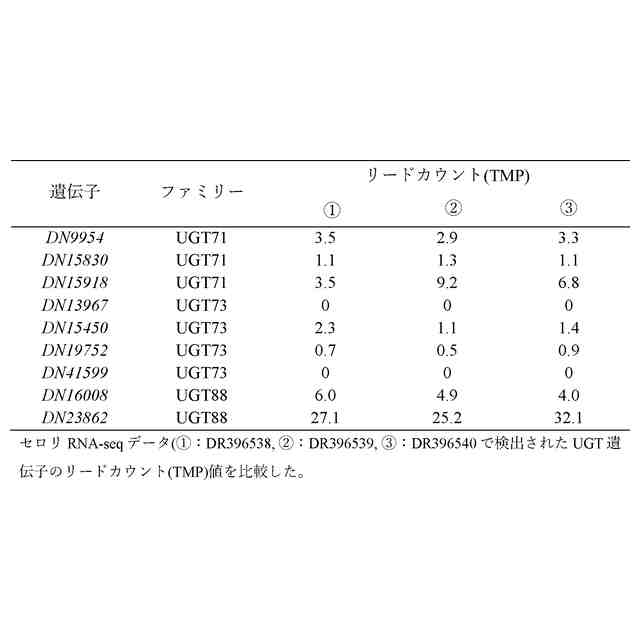
AgGlcT候補遺伝子とUGTの系統樹を示す図である。セロリRNA-seqのリードデータ(SRR1023730)から発現量の高いUGT71、72、73、78、88のホモログを検索した。糖の結合位置に基づいてクラスターを分けた。丸は9つの候補遺伝子(セロリゲノムにコードされるUGT71、73、88)、四角は7つの候補遺伝子(セロリ葉RNA-seqで発現が見られた遺伝子)、星は3つの候補遺伝子(アピイン生合成酵素遺伝子AgFNS IとAgApiTと発現挙動が比較的類似していた遺伝子)を示す。
AgGlcT候補遺伝子セロリ発達段階ごとの半定量PCRを示す図である。(A)アピイン生合成関連酵素遺伝子(FNS I、ApiT)と7つの候補遺伝子の発達段階ごとの遺伝子発現量を半定量PCRで定量した。GAPDHは内部標準として用いた。(B)アピイン生合成関連酵素遺伝子(FNS I、ApiT)と7つの候補遺伝子の発達段階ごとの遺伝子発現量を半定量PCRで定量した結果を示すグラフ。
セロリ本葉発達段階ごとのアピイン生合成関連酵素遺伝子(AgGlcT, AgApiT, AgFNS I)の発現量を示す図である。(A)セロリ本葉発達段階ごとのAgGlcTの発現量。(B)セロリ本葉発達段階ごとのAgApiTの発現量。(C)セロリ本葉発達段階ごとのAgFNS Iの発現量。0.5 cmにおける各遺伝子の発現量を1とした。
AgGlcT候補タンパク質の発現を示す図である。
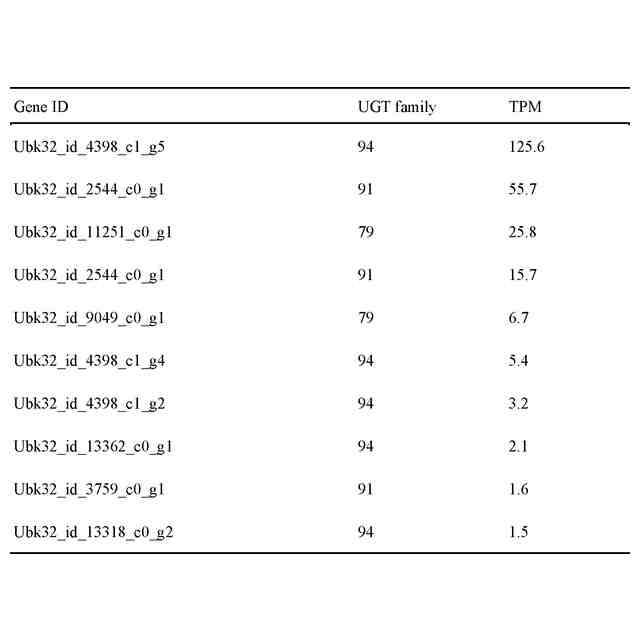
AgGlcT候補タンパク質の酵素活性を示す図である。(A)生成物の定量はHPCL<条件1>で行った。上からDN23862、DN16008、DN9954のピークを示す。▼はアピゲニン7-O-グルコシド、▽はアピゲニンのピークを示す。(B)生成物の酵素活性を示す。
AgGlcTの酵素量依存性・時間依存性を示す図である。(A)1 mMアピゲニン、0.5 mM UDP-Glc、パセリ素酵素液1 μgをpH7.0、23℃、0~4時間で反応させた。生成物の定量はHPCL<条件1>で行った。(B)1 mMアピゲニン、0.5 mM UDP-Glc、セロリ素酵素液10~1.8 μgをpH7.0、23℃、2時間で反応させた。生成物の定量はHPLC<条件1>で行った。
AgGlcTのアミノ酸配列アライメントを示す図である。下線は触媒残基、斜体はPSPGモチーフ、□はGSSモチーフを示す。
AgGlcTの立体構造予測を示す図である。
AgGlcTのpH依存性を示す図である。
AgGlcTのドナー基質特異性を示す図である。(A)生成物の定量はHPCL<条件1>で行った。(B)酵素活性が検出された上位2つの基質の構造を示す。AgGlcTはUDP-Glcの丸で示した部分を認識している。
AgGlcTのアクセプター基質依存性を示す図である。(A)生成物の定量はHPCL<条件1>で行った。(B)酵素活性が検出された6つの基質の構造を示す。AgGlcTはアピゲニン、ケルセチン、ルテオリン、クリソエリオールの丸で示した部分を認識している。
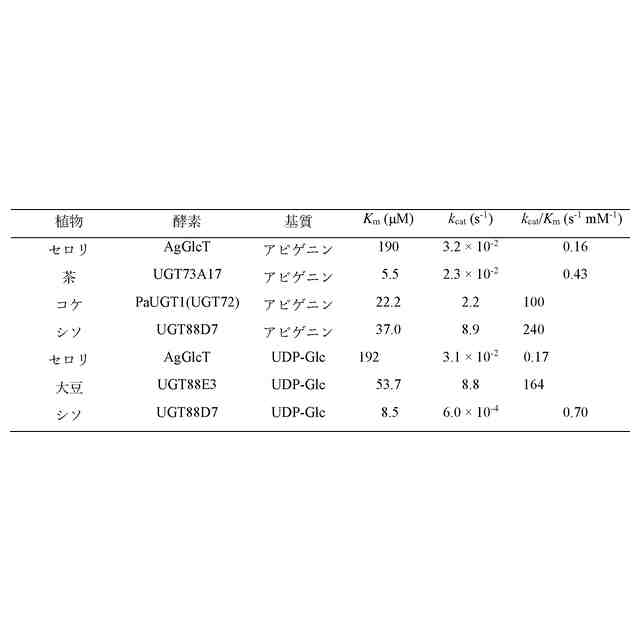
AgGlcTの反応速度論的解析を示す図である。(A)UDP-Glcに対する反応速度を求めた。UDP-Glcは0~500 μMの複数の濃度を用いた。K
m
= 192.2 μM、V
max
= 36.2 p mol/min/μgであった。(B) アピゲニンに対する反応速度を求めた。アピゲニンは0~1,000 μMまでの複数の濃度を用いた。K
m
= 190.0 μM、V
max
= 37.6 p mol/min/μgであった。
PcGlcTとAgGlcT、PcApiTとAgApiTのアミノ酸配列アライメントを示す図である。
(A) 組換えPcGlcTタンパク質のための発現コンストラクトを示す図である。 (B) 精製した組換えPcGlcTタンパク質のSDS-PAGEを示す図である。(C) 組換えPcApiTタンパク質のための発現コンストラクトを示す図である。 (D) 精製した組換えProS2-PcApiT融合タンパク質とタグ切断したPcApiTタンパク質のSDS-PAGEを示す図である。
(A) PcGlcTとPcApiTが触媒するアピイン生合成を示す図である。(B) PcGlcTの酵素反応のHPLC分析を示す図である。(C) PcApiTの酵素反応のHPLC分析を示す図である。
(A) PcGlcTのアクセプター基質特異性を示す図である。(B) PcGlcTのドナー基質特異性を示す図である。 (C) PcApiTのアクセプター基質特異性を示す図である。(D) PcApiTのドナー基質特異性を示す図である。 3回測定した平均と標準偏差を示している
(A) PcGlcTの至適pHを示す図である。(B) PcApiTの至適pHを示す図である。
(A) qRT-PCRによる PcFNSIの各器官の発現パターンと本葉の発達段階での発現量を示す図である。 (B) qRT-PCRによる PcGlcTの各器官の発現パターンと本葉の発達段階での発現量を示す図である。 (C) qRT-PCRによるPcApiTの各器官の発現パターンと本葉の発達段階での発現量を示す図である。
PcGlcTとPcApiTの遺伝子塩基配列(コーディング領域のみ)とタンパク質アミノ酸配列を示す図である。
API001(左)およびAPI002(右)の可溶性画分のSDS-PAGEおよびウエスタンブロット解析の結果を示す図である。上段:CBB染色、中段:Anti-Hisタグを用いたウエスタンブロッティング、下段:Anti-Sタグを用いたウエスタンブロッティング。
API001およびAPI002における逆相HPLCの結果を示す図である。NC:pColdProS2、pCDF-1b及びpACYCDuetのみが導入された大腸菌株。Apiin:アピイン標品のピーク位置。A7G:A7G標品のピーク位置。Apigenin:アピゲニン標品のピーク位置。
API001及びAPI002におけるA7G及びアピインの生産量を示す図である。
【発明を実施するための形態】
【0010】
1.アピオース転移酵素およびグルコース転移酵素
本発明は、アピオース転移酵素(以下、本発明のアピオース転移酵素)を提供する。本発明はまた、グルコース転移酵素(以下、本発明のグルコース転移酵素)を提供する。
(【0011】以降は省略されています)
この特許をJ-PlatPat(特許庁公式サイト)で参照する
関連特許
学校法人立命館
光演算装置
6か月前
学校法人立命館
味覚向上装置
12か月前
学校法人立命館
液面レベルセンサ
3か月前
学校法人立命館
情報埋め込み方法
4か月前
学校法人立命館
バイオ炭の分析方法
3か月前
学校法人立命館
センサ及びセンサシステム
3か月前
学校法人立命館
流路構造、流路構造製造方法
7か月前
学校法人立命館
運動誘導装置、運動誘導方法
8か月前
学校法人立命館
ロボットハンド制御システム
1か月前
学校法人立命館
位置同定システム及びその方法
5日前
トヨタ自動車株式会社
色処理方法
9か月前
学校法人立命館
周波数シンセサイザ及び無線通信装置
11か月前
株式会社東洋レーベル
入眠促進装置
6か月前
株式会社豊田自動織機
電力変換装置
2か月前
学校法人立命館
コマンド入力装置、コマンド入力方法
9か月前
三洋化成工業株式会社
金属回収方法
5か月前
学校法人立命館
配管パイプ型微生物燃料電池及び配管システム
11か月前
学校法人立命館
筋組織カルシウム蓄積を抑制するビタミンD作用
7か月前
学校法人立命館
フェーズドアレーアンテナ装置及び無線通信装置
5か月前
学校法人立命館
個人認証装置、個人認証方法、個人認証プログラム
4か月前
学校法人立命館
感情推定装置、感情推定方法、感情推定プログラム
9か月前
Patentix株式会社
霧化装置及び製膜装置。
8か月前
白山工業株式会社
液体タンクの床板検査用ロボット
5日前
学校法人立命館
処理装置、音響システム、先行音効果を生じさせる方法
2か月前
学校法人立命館
異種タンパク質の大量生産が可能なナス科植物の四重変異体
6か月前
兵庫県
強化炭素繊維、炭素繊維シート及び炭素繊維強化剤
2か月前
学校法人立命館
薄膜の膜厚を測定する測定方法、測定装置および測定プログラム
9か月前
学校法人立命館
設計システム、コンピュータ実装方法、及びコンピュータプログラム
9か月前
学校法人立命館
心理的距離推定装置、心理的距離推定方法、心理的距離推定プログラム
10か月前
学校法人立命館
アピイン生産のための新規遺伝子およびそれを用いたアピインの生産方法
10か月前
学校法人立命館
メッシュ生成システム、コンピュータ実装方法及びコンピュータプログラム
7か月前
Patentix株式会社
積層構造体、半導体装置、電子機器及びシステム
8か月前
Patentix株式会社
積層構造体、半導体装置、電子機器及びシステム
8か月前
Patentix株式会社
積層構造体、半導体装置、電子機器及びシステム
8か月前
日工株式会社
モルタルの製造方法およびフレッシュコンクリートの製造方法
11か月前
Patentix株式会社
積層構造体、半導体装置、電子機器及びシステム
8か月前
続きを見る
他の特許を見る




 特許ウォッチ
特許ウォッチ