TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2024128959
公報種別
公開特許公報(A)
公開日
2024-09-24
出願番号
2024034653
出願日
2024-03-07
発明の名称
コロナウイルス感染症治療薬のスクリーニング方法
出願人
国立研究開発法人理化学研究所
,
個人
代理人
弁理士法人特許事務所サイクス
主分類
C12N
15/113 20100101AFI20240913BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】 コロナウイルスのRNA依存性RNAポリメラーゼまたはプライマーゼが結合する二次構造を形成する塩基配列を含むRNA、およびこのRNAを用いたコロナウイルス感染症の治療剤または予防剤のスクリーニング方法を提供すること。
【解決手段】 シュードノット構造を形成する塩基配列を含むRNA、およびこのRNAを用いたコロナウイルス感染症の治療剤または予防剤のスクリーニング方法が提供される。
【選択図】図7
特許請求の範囲
【請求項1】
配列番号22に記載の塩基配列、または
配列番号22に記載の塩基配列と少なくとも90%の塩基配列同一性を有し且つシュードノット構造を形成する塩基配列を含む、RNA。
続きを表示(約 880 文字)
【請求項2】
配列番号21に記載の塩基配列、または配列番号21に記載の塩基配列と少なくとも90%の塩基配列同一性を有し且つ3つのステム構造を形成する塩基配列を含む、RNA。
【請求項3】
配列番号20に記載の塩基配列、または配列番号20に記載の塩基配列と少なくとも90%の塩基配列同一性を有し且つシュードノット構造を形成する塩基配列を含む、RNA。
【請求項4】
配列番号19に記載の塩基配列、または配列番号19に記載の塩基配列と少なくとも90%の塩基配列同一性を有し且つ2つのステム構造を形成する塩基配列を含む、RNA。
【請求項5】
シュードノット構造に加えて1つのステムループ構造を形成する、請求項1に記載のRNA。
【請求項6】
請求項1に記載のRNA、並びに
請求項2から4のいずれか一項に記載のRNAから選択される1種、2種、または3種のRNAを含む、セット。
【請求項7】
請求項1に記載のRNAとプライマーゼ/RNA依存性RNAポリメラーゼ(RdRp)との結合を検出するアッセイを使用して、プライマーゼ/RdRpのRNAへの結合を阻害する化合物を同定する工程を含む、
コロナウイルス感染症の治療剤または予防剤のスクリーニング方法。
【請求項8】
核磁気共鳴(NMR)分析により、請求項1に記載のRNAについて化合物の結合部位を解析する工程をさらに含む、請求項7に記載のスクリーニング方法。
【請求項9】
同定された化合物の存在下で、請求項2から4のいずれか一項に記載のRNAとプライマーゼ/RNA依存性RNAポリメラーゼ(RdRp)との結合を分析する工程をさらに含む、請求項7に記載のスクリーニング方法。
【請求項10】
コロナウイルスがアルファコロナウイルス属またはベータコロナウイルス属のウイルスである、請求項7に記載のスクリーニング方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、シュードノット構造を形成する塩基配列を含むRNAに関する。本発明はさらに、このRNAを用いたコロナウイルス感染症の治療剤または予防剤のスクリーニング方法に関する。
続きを表示(約 8,100 文字)
【背景技術】
【0002】
日本で承認されている新型コロナウイルス感染症(COVID-19)の治療薬のうち、レムデシビルとモルヌピラビルはRNA合成阻害剤である(非特許文献1)。レムデシビルはアデノシンヌクレオシドのプロドラッグである。生体内で加水分解などを経て生成される活性代謝物であるアデノシン三リン酸(ATP)の類似体が、RNAウイルスの複製に必要なRNA依存性RNAポリメラーゼを阻害することで抗ウイルス作用を発揮する(非特許文献1)。モルヌピラビルもプロドラッグであり、N-ヒドロキシシチジン(NHC)に代謝され細胞内に取り込まれた後、活性型であるリボヌクレオシド三リン酸化体(NHC-TP)にリン酸化され、NHC-TPがウイルス由来RNA依存性RNAポリメラーゼによりウイルスRNAに取り込まれた結果、ウイルスゲノムのエラー頻度が増加し、ウイルスの増殖が阻害される(非特許文献2)。すなわちこれら2剤はRNA依存性RNAポリメラーゼ(RdRp)阻害剤である。RdRpは、RNAウイルスの複製過程を担う重要な酵素であり、COVID-19治療薬の開発の有用な標的の一つである。しかしながら、両剤ともCOVID-19に対する治療効果が十分であるとはいえないものである。
【0003】
コロナウイルスはプラス鎖一本鎖のRNAをウイルスゲノムとして有し、その3′末端非翻訳領域(UTR)から(-)鎖ゲノムRNAとサブゲノムRNAが転写される。非特許文献3は、コロナウイルス(-)鎖RNA合成の開始に関する仮説的モデルを開示する。この仮説的モデルでは、プライマーゼがシュードノットのループ(ループ1)になる部分と3′末端が形成するステムに結合してプライマーゼが(-)鎖RNAの5′末端を合成することで、ゲノムの3′末端がループ1から離れてシュードノットが形成され、このシュードノットにRdRpが結合して(-)鎖が伸長することが提唱されている。しかしながら、コロナウイルスゲノムの3′末端においてシュードノット構造が形成されるかについては実験的に確認されてなかったため、コロナウイルス(-)鎖RNAの合成開始機構の詳細は分かっていなかった。
【先行技術文献】
【非特許文献】
【0004】
Ghosh, Arun K et al. “Recent Drug Development and Medicinal Chemistry Approaches for the Treatment of SARS-CoV-2 Infection and COVID-19.” ChemMedChem vol. 17,22 (2022): e202200440. doi:10.1002/cmdc.202200440.
Imran M, Kumar Arora M, Asdaq SMB, Khan SA, Alaqel SI, Alshammari MK, Alshehri MM, Alshrari AS, Mateq Ali A, Al-Shammeri AM, Alhazmi BD, Harshan AA, Alam MT, Abida. Discovery, Development, and Patent Trends on Molnupiravir: A Prospective Oral Treatment for COVID-19. Molecules. 2021 Sep 24;26(19):5795. doi: 10.3390/molecules26195795. PMID: 34641339; PMCID: PMC8510125.
Zust, R., Miller, T. B., Goebel, S. J., Thiel, V. & Masters, P. S. Genetic interactions between an essential 3' cis-acting RNA pseudoknot, replicase gene products, and the extreme 3' end of the mouse coronavirus genome. J Virol 82, 1214-1228 (2008). https://doi.org:10.1128/JVI.01690-07
【発明の概要】
【発明が解決しようとする課題】
【0005】
上記のとおり、RdRpは、RNAウイルスの複製過程を担う重要な酵素であるため、これを標的とするCOVID-19治療薬の開発が求められている。コロナウイルスのRdRp阻害剤の開発には、(-)鎖gRNAの転写開始点であり特異な構造を形成するウイルスゲノムの3′末端の二次構造の詳細を実験的に明らかにし、それをin vitroで再構築する必要がある。
【0006】
よって、本発明の課題は、コロナウイルスのRdRpまたはプライマーゼが結合する二次構造を形成する塩基配列を含むRNAを提供すること、およびこのRNAを用いたコロナウイルス感染症の治療剤または予防剤のスクリーニング方法を提供することである。
【課題を解決するための手段】
【0007】
本発明者らは、SARS-CoV-2の3′UTRの塩基配列を基にRNAコンストラクトを設計作成し、3′UTRの3′PKと呼ばれている部分がステムループ構造とシュードノット構造を相互に排他的にとることをNMRにより初めて原子レベルで証明した。P1pkとP2領域によるシュードノット形成に伴い、ステムループ構造から抜け出し、3′末端の配列を含むP5ステムが開口することを明らかにした。ゲルシフトアッセイにより、nsp7とnsp8の1対1混合物は、ステムループとシュードノットの両方のコンフォメーションと約2μMの見かけの親和性で相互作用することが明らかになった。NMR解析の結果、nsp7とnsp8の相互作用はステムループ構造においてはP5ステムを不安定化させ、シュードノット型は変化させないことがわかった。これらの結果は、シュードノットとnsp7-nsp8複合体の相互作用により、ウイルスgRNAの3′末端がRNA合成に適した一本鎖RNAに変換されることを示唆しており、シュードノットが抗ウイルス薬開発の有用な標的であることを示している。本発明はこの知見に基づくものである。
【0008】
本発明によれば以下の発明が提供される。
[1] 配列番号22に記載の塩基配列、または配列番号22に記載の塩基配列と少なくとも90%の塩基配列同一性を有し且つシュードノット構造を形成する塩基配列を含む、RNA。
[2] 配列番号21に記載の塩基配列、または配列番号21に記載の塩基配列と少なくとも90%の塩基配列同一性を有し且つ3つのステム構造を形成する塩基配列を含む、RNA。
[3] 配列番号20に記載の塩基配列、または配列番号20に記載の塩基配列と少なくとも90%の塩基配列同一性を有し且つシュードノット構造を形成する塩基配列を含む、RNA。
[4] 配列番号19に記載の塩基配列、または配列番号19に記載の塩基配列と少なくとも90%の塩基配列同一性を有し且つ2つのステム構造を形成する塩基配列を含む、RNA。
[5] シュードノット構造に加えて、1つのステムループ構造を形成する、[1]に記載のRNA。
[6] [1]に記載のRNA、並びに[2]から[4]のいずれか一に記載のRNAから選択される1種、2種、または3種のRNAを含む、セット。
[7] [1]に記載のRNAとプライマーゼ/RNA依存性RNAポリメラーゼ(RdRp)との結合を検出するアッセイを使用して、プライマーゼ/RdRpのRNAへの結合を阻害する化合物を同定する工程を含む、コロナウイルス感染症の治療剤または予防剤のスクリーニング方法。
[8] 核磁気共鳴(NMR)分析により、[1]に記載のRNAについて化合物の結合部位を解析する工程をさらに含む、[7]に記載のスクリーニング方法。
[9] 同定された化合物の存在下で、[2]から[4]のいずれか一に記載のRNAとプライマーゼ/RNA依存性RNAポリメラーゼ(RdRp)との結合を分析する工程をさらに含む、[7]に記載のスクリーニング方法。
[10] コロナウイルスがアルファコロナウイルス属またはベータコロナウイルス属のウイルスである、[7]に記載のスクリーニング方法。
[11] コロナウイルスがSARS-CoV-2である、[7]に記載のスクリーニング方法。
[12] [1]から[4]のいずれか一に記載のRNAをコードするDNA。
[13] [1]から[4]のいずれか一に記載のRNAについてNMR解析により二次構造の形成を測定する方法。
[14] プライマーゼまたはRdRpのRNAにおける結合部位を特定するための、[13]に記載の測定方法。
[15] [1]から[4]のいずれか一にRNAをアッセイ溶液中で維持することを含む、シュードノット構造またはステムループ構造の形成方法であって、アッセイ溶液のカリウムイオン濃度がナトリウムイオン濃度よりも高い、方法。
[16] [1]から[4]のいずれか一に記載のRNAの少なくとも一部分に結合して、前記RNAとプライマーゼ/RNA依存性RNAポリメラーゼ(RdRp)との結合を阻害する、アンチセンスオリゴヌクレオチド。
[17] 下記(a)~(o)からなる群から選択される何れかの塩基配列(但し、ウラシルはチミンであってもよい)を含む、[16]に記載のアンチセンスオリゴヌクレオチド:
(a)GGGAAUAGCUUCUUAGGAGAAU(配列番号25)、
(b)AAUAGCUUCUUAGGAGAAU(配列番号27)、
(c)GAUUAAAGUUAAUUACGAGAAUU(配列番号28)、
(d)AAAGUCGAGAAUUCAUUCAUUCU(配列番号29)、
(e)UUGUGCUAUGUAGUUACGAGA(配列番号30)、
(f)UGUGAGAUUAAAGUUAACUACAUCUAC(配列番号31)、
(g)UUGUGCUAUGUAGUUA(配列番号32)、
(h)UGUGAGAUUAAAGUUAACUACA(配列番号33)、
(i)AUUCUCCUAAGAAGCUAUU(配列番号34)、
(j)UAUGUAGUUACGAGAAUUCAUUCU(配列番号35)、
(k)AAAGUCGAGAAUUCAUUCAUUCUGCACAAG(配列番号36)、
(l)AUCUACUUGUGCUAUGUAGUUACGAGAA(配列番号37)、
(m)UGUGAGAUUAAAGUUAACUACAUCUAU(配列番号38)、
(n)(a)~(m)の塩基配列において1から数個の塩基が欠失、置換、挿入又は付加塩基配列された塩基配列から成り、[1]から[4]のいずれか一に記載のRNAとRdRpとの結合を阻害することができる、塩基配列、および
(o)(a)~(m)のいずれか一の塩基配列と90%塩基配列同一性を有する配列を含み、[1]から[4]のいずれか一に記載のRNAとRdRpとの結合を阻害することができる、塩基配列。
【図面の簡単な説明】
【0009】
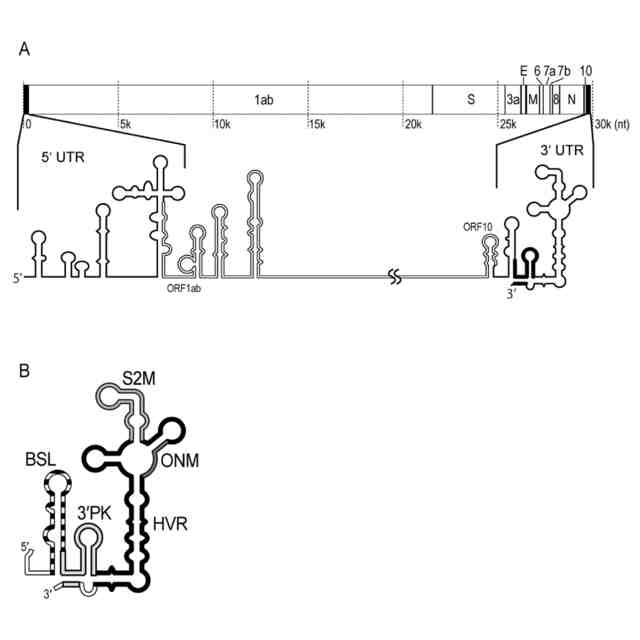
(A) SARS-CoV-2のゲノムRNAと二次構造の模式図。(B)3′PK領域の二次構造の模式図。BSL:バルジドステムループ、HVR:超可変領域、ONM:オクタヌクレオチドモチーフ、S2M:ステムループII様モチーフ、3′PK:BSLの一部とBSLとHVR構造の間のステムを含む領域。
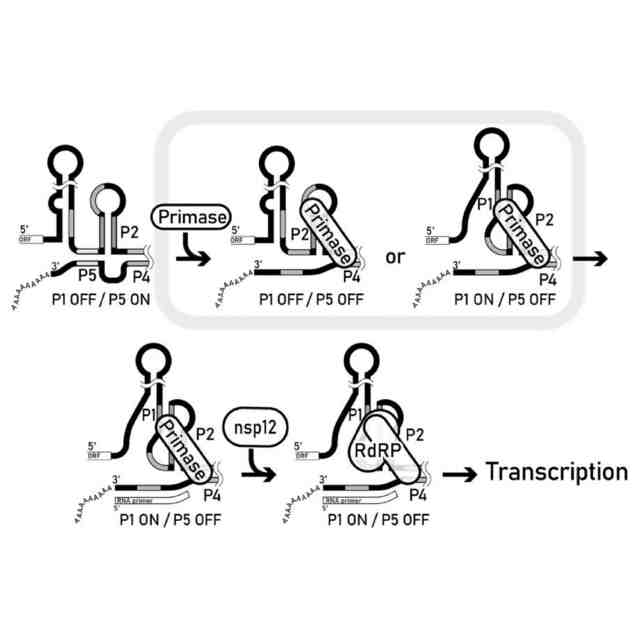
Zustら(非特許文献3)の提案に基づき、若干の修正を加えたコロナウイルスの仮想的なゲノムRNA複製開始モデル。(A)3′PKにステムループ構造が形成される。(B)nsp7、nsp8、nsp9からなるプライマーゼがP5ステムに結合する。(C)プライマーゼは(-)鎖gRNAのプライマーを転写する。(D)プライマーがgRNAの3′末端にプライミングされ、P5ステムが対になっていないため、シュードノット構造が形成される。(E)Nsp12がシュードノット構造とnsp7-nsp8-nsp9複合体に結合し、(-)鎖gRNAを伸長させる。
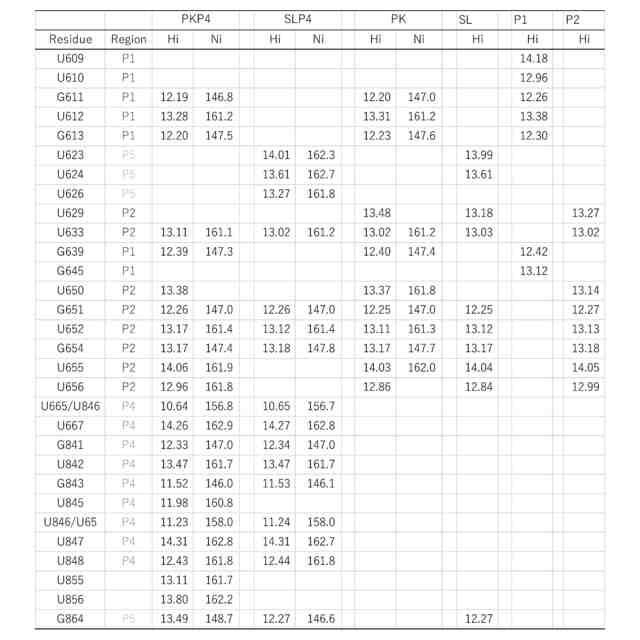
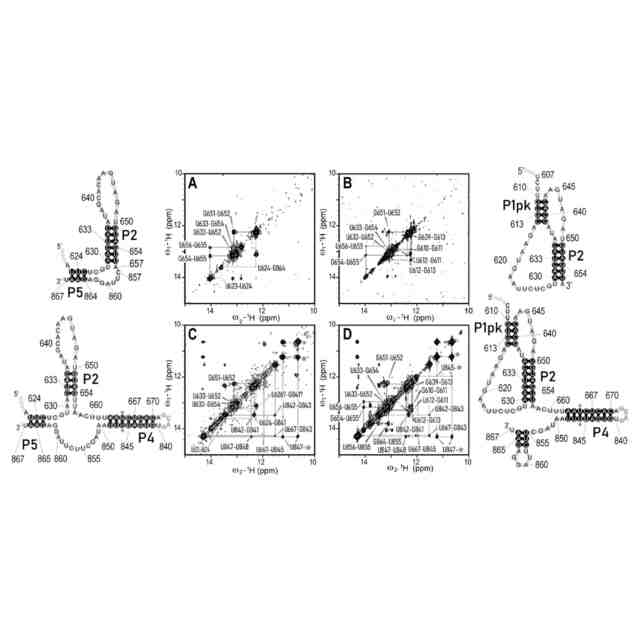
(A)SL、(B)PK、(C)SLP4、(D)PKP4の3′PK断片の2D NOESYスペクトルのイミノプロトン領域と、NMRに基づく対応する二次構造との比較。図では簡略化のため、塩基番号の最初の2桁(29XXX)は省略した。塗りつぶした丸で示したNOEはG639-UおそらくU637またはU650である。アスタリスクで示したクロスピークはU665とU846イミノプロトン間の塩基対内NOEである。U845-★はU845とU665またはU846間のNOE、U847-★はU847とU665またはU846間のNOE。
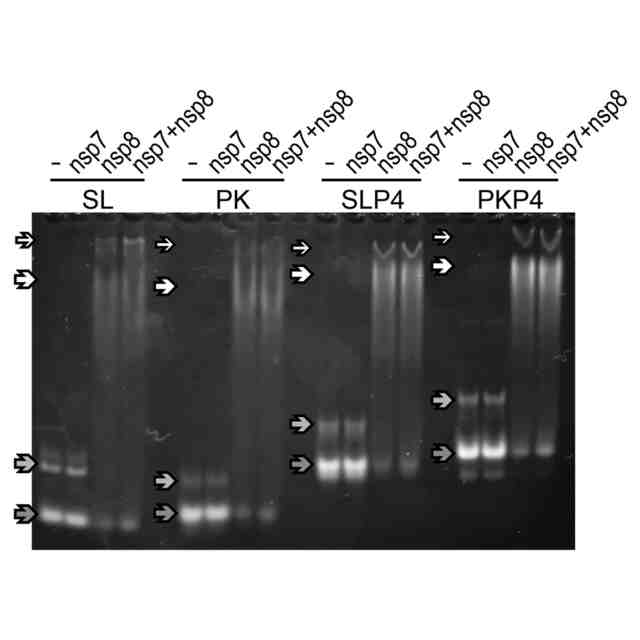
nsp7およびnsp8との結合について試験した4つのRNA断片(SL、PK、SLP4、PKP4)のゲル画像。一番下の矢印(ウエルから最も離れたもの)で示したバンドは構造を形成したRNA、下から二番目の矢印で示したバンドは構造を形成していない(直鎖状)RNAに対応し、上から二番目と一番目の矢印で示したバンドはそれぞれタンパク質と結合したRNA、タンパク質と結合したRNAの別のコンフォーマーに対応する。RNA断片は、nsp8またはnsp7-nsp8混合物の添加により、バンドがシフトすることがわかった。
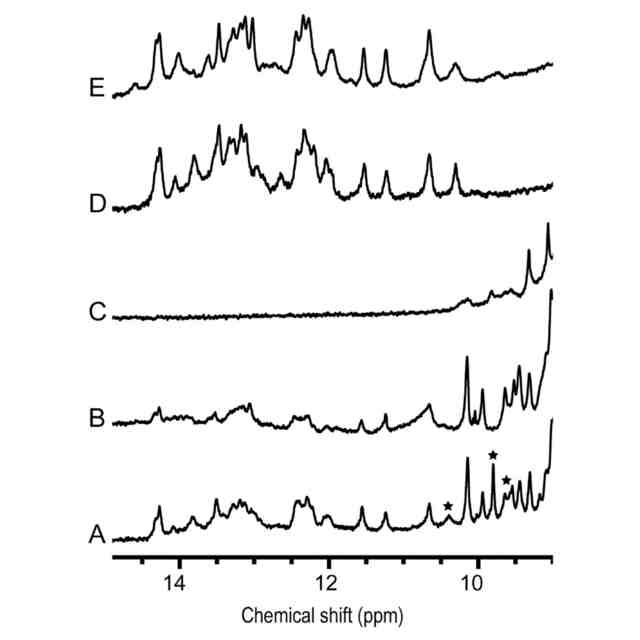
RNAとnsp7-nsp8混合物の複合体形成による構造変化。(A)PKP4,nsp7,nsp8の1:2:2混合物,(B)SLP4,nsp7,nsp8の1:1:1混合物,(C)nsp7とnsp8の1:1混合物,(D)PKP4,および(E)SLP4の1D
1
H NMRスペクトルのイミノプロトン領域。星印で示したタンパク質によるシグナルは、PKP4-nsp7-nsp8混合物でのみ観察された。
nsp7-nsp8混合物の結合によるSLP4とPKP4の構造変化。
1
H-
1
H 2D NOESY スペクトルのイミノプロトン領域(A)SLP4,(B)PKP4,(C)SLP4,nsp7,nsp8の1:1:1混合物および(D)PKP4,nsp7,nsp8の1:2:2混合物。
nsp7-nsp8混合物の結合によるSLP4とPKP4の構造変化。(E)nsp7-nsp8とSLP4の結合の模式図。SLP4に存在するP5ステム領域の解離は、塩基対内NOEシグナルの欠如によって確認された。(F)nsp7-nsp8と結合したPKP4の二次構造の模式図。nsp7-nsp8との結合による化学シフトの変化Δδを示す。
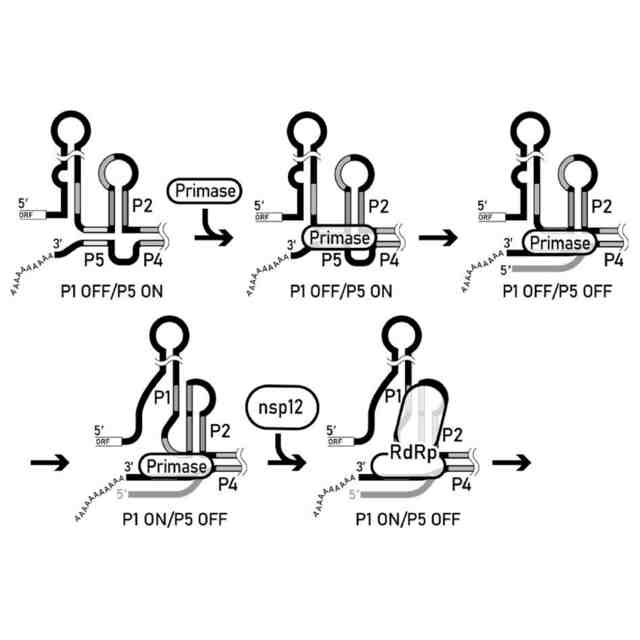
コロナウイルスの改変(-)鎖ゲノムRNA複製開始仮説モデル。本発明者らにより提案されるモデルである。(A)3′PKにステムループ構造が形成される。(B)nsp7-nsp8複合体(図中ではPrimaseと記載)がステムループ構造に結合するか、(C)シュードノット構造に直接結合し、P5が一本鎖に変換される。(D)プライマーゼによりRNAプライマーが合成される。(E)Nsp12がシュードノット構造およびプライマーゼに結合し、(-)鎖gRNAを伸長させる。
P1の2D NOESYスペクトルのイミノプロトン領域と、NMRに基づく対応する二次構造との比較。図では簡略化のため、塩基番号の最初の2桁(29XXX)は省略した。
P2の2D NOESYスペクトルのイミノプロトン領域と、NMRに基づく対応する二次構造との比較。図では簡略化のため、塩基番号の最初の2桁(29XXX)は省略した。
(A)SL,(B)SLL,(C)SLP4のイミノプロトン領域。(D)SL(E)SLL,(F)SLP4のNMRによる二次構造の模式図。SLLとSLP4のU29655とU29656(P2ステムに含まれる)に起因するシグナルは、SLに対して著しく低い強度を示した。
(A)PK、SL、SLP4、PKP4のゲルシフトアッセイ。下から(移動度の早い)順に矢印で、タンパク質と結合していないが構造を形成しているRNA、タンパク質と結合していない直鎖状RNA、RNA-nsp7-nsp8複合体を示した。(B)各RNA断片のHill-Langmuirプロット。PKのKdは5.1μM、SLは5.5μM、PKP4は2.5μM、SLP4は2.3μMであった。パラメータθ=1-([タンパク質と結合していないRNA]/[RNA総量]),[タンパク質と結合していないRNA]/[RNA総量]はタンパク質とRNAを混合したサンプルの各タンパク質と結合していないが構造を形成しているRNA のバンドとRNAのみの構造を形成しているRNAのバンドの体積比から求めた。P4とL4/5を持つ断片は、他の断片よりも親和性が高いことがわかった。また、全ての断片がn>1を示し、nsp7-nsp8が正の協同性を示すことが示された。
(A)nsp7-nsp8、(B)PKP4-nsp7-nsp8混合物、(C)SLP4-nsp7-nsp8混合物の
1
H-
1
H 2D NOESYスペクトルのタンパク質アミドプロトン領域。PKP4やSLP4の添加により、RNA-タンパク質複合体による新たなNOEシグナルが多数観測された。RNA-タンパク質複合体のNOEパターンは、結合するRNAの構造によって部分的に異なることがわかった。
実施例4のアンチセンスRNAを用いた実験の電気泳動のゲル写真である。
実施例5のアンチセンスRNAを用いた実験の電気泳動のゲル写真である。
アンチセンスRNA#2(配列番号28)、#3(配列番号29)、#4(配列番号30)および#5(配列番号31)のSLP4およびPKP4への結合位置を示す概要図である。
アンチセンスRNA#8(配列番号34)、#9(配列番号35)、#10(配列番号36)および#11(配列番号37)のSLP4およびPKP4への結合位置を示す概要図である。
実施例6のアンチセンスRNAを用いた実験の電気泳動のゲル写真である。
実施例6における、結合していないRNAのバンドボリューム比を示すグラフである。
【発明を実施するための形態】
【0010】
以下に記載する本発明の説明は、代表的な実施形態や具体例に基づいてなされることがあるが、本発明はそのような実施形態に限定されるものではない。なお、本明細書において「~」を用いて表される数値範囲は「~」の前後に記載される数値を下限値および上限値として含む範囲を意味する。
(【0011】以降は省略されています)
特許ウォッチbot のツイートを見る
この特許をJ-PlatPatで参照する
関連特許
マグネデザイン株式会社
磁気顕微鏡
1か月前
東洋紡株式会社
細菌からの核酸抽出法
3日前
個人
セルロース性物質の製造方法
2か月前
株式会社ゴーフォトン
PCR方法
1か月前
熊本県
低褐変レタスとその作製方法
11日前
東洋紡株式会社
ウイルスからの核酸抽出法
3日前
SMC株式会社
気体供給装置
1か月前
朝日酒造 株式会社
発泡性清酒の製造方法
今日
テルモ株式会社
液体除去器具
28日前
国立大学法人山梨大学
受精胚の選別方法及び装置
22日前
鹿島建設株式会社
褐藻の冷凍保存方法
1か月前
サッポロビール株式会社
アルコール飲料
2か月前
株式会社テクノーブル
乳酸菌及び皮膚外用剤
1か月前
サッポロビール株式会社
アルコール飲料
2か月前
サッポロビール株式会社
アルコール飲料
1か月前
アサヒビール株式会社
容器詰麦芽発酵飲料
2か月前
学校法人君が淵学園
核酸の部位特異的アシル修飾剤
2か月前
株式会社SUPER BLOOM
スピリッツ
1か月前
テルモ株式会社
移植用デバイス
28日前
新東工業株式会社
培養システム
1か月前
新東工業株式会社
培養システム
1か月前
テルモ株式会社
積層体および移植片の輸送方法
28日前
ヤマサ醤油株式会社
アルギニン非分解性乳酸菌株の取得方法
2か月前
サッポロビール株式会社
茶風味アルコール飲料
1か月前
株式会社ユーグレナ
観察用遠隔制御装置
28日前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
2か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
2か月前
住友化学株式会社
細胞用基材
22日前
住友化学株式会社
細胞用基材
22日前
日本臓器製薬株式会社
髄核細胞の培養方法
21日前
サムコ株式会社
ポアデバイス
2か月前
三井化学株式会社
作業用装置
22日前
株式会社関電工
藻類培養装置および藻類培養方法
1か月前
株式会社アステック
培養装置及び培養方法
1か月前
TOA株式会社
化粧品の保存効力試験方法
28日前
学校法人慈恵大学
培養皿用の定規
今日
続きを見る
他の特許を見る
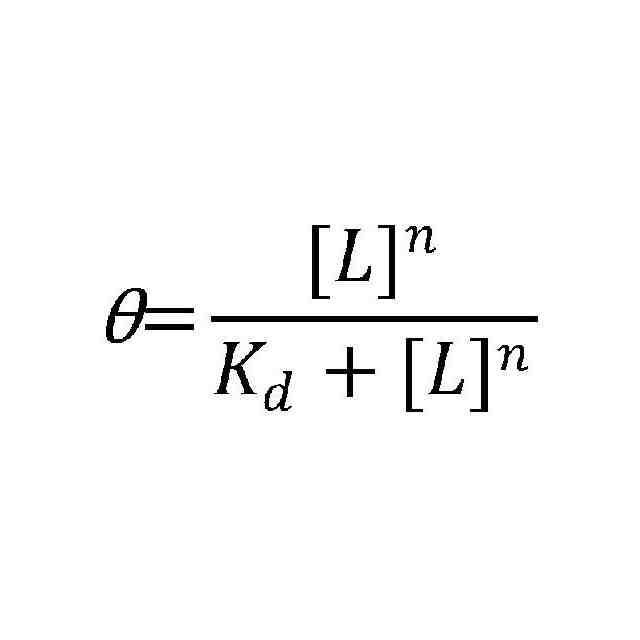

 特許ウォッチ
特許ウォッチ