TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2024125388
公報種別
公開特許公報(A)
公開日
2024-09-18
出願番号
2024104650,2022089545
出願日
2024-06-28,2010-10-25
発明の名称
グリコシル化された免疫グロブリンの調製方法
出願人
エフ. ホフマン-ラ ロシュ アーゲー
,
F. HOFFMANN-LA ROCHE AKTIENGESELLSCHAFT
,
中外製薬株式会社
代理人
個人
,
個人
,
個人
,
個人
主分類
C12P
21/08 20060101AFI20240910BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】調製される免疫グロブリンのグリコシル化パターンを培養条件に基づいて改変できる、免疫グロブリンの調製方法を提供する。
【解決手段】a)免疫グロブリンをコードする核酸を含む真核細胞を用意し、b)培地中の単位時間当たり利用できるグルコースの量が一定に維持され、かつ単位時間当たりその培地中でその細胞が最大限に利用できる量の80%未満に制限された培地中で、前記の真核細胞を培養し、そしてc)培養物から免疫グロブリンを回収する。
【選択図】図1
特許請求の範囲
【請求項1】
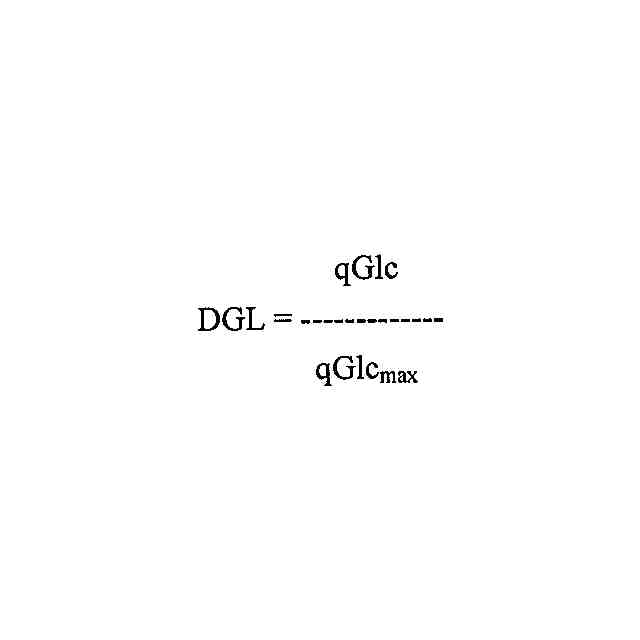
a)免疫グロブリンをコードする核酸を含む真核細胞を、培地中での単一細胞の現時点のグルコース比消費速度-対-その種類の細胞の既知の最大グルコース比消費速度の比が一定に維持された培地中で培養し、そして
b)培養物から免疫グロブリンを回収する
ことを含む、免疫グロブリンの調製方法。
続きを表示(約 700 文字)
【請求項2】
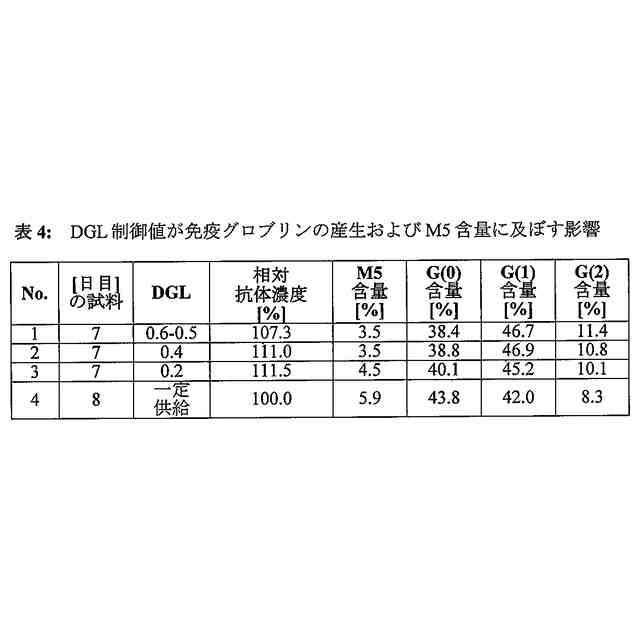
比が0.8から0.2までであることを特徴とする、請求項1に記載の方法。
【請求項3】
比が0.6から0.4までであることを特徴とする、請求項2に記載の方法。
【請求項4】
a)免疫グロブリンをコードする核酸を含む真核細胞を、培地中の単位時間当たり利用できるグルコースの量が一定に維持され、かつ単位時間当たりその培地中でその細胞が最大限に利用できる量の80%未満の一定値に制限された培地中で培養し、そして
b)培養物から免疫グロブリンを回収する
ことを含む、免疫グロブリンの調製方法。
【請求項5】
一定値が80%未満であって20%を超えることを特徴とする、請求項4に記載の方法。
【請求項6】
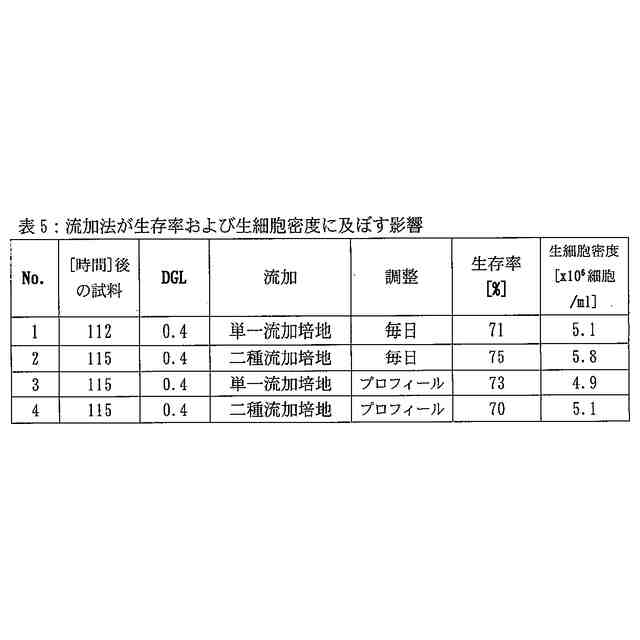
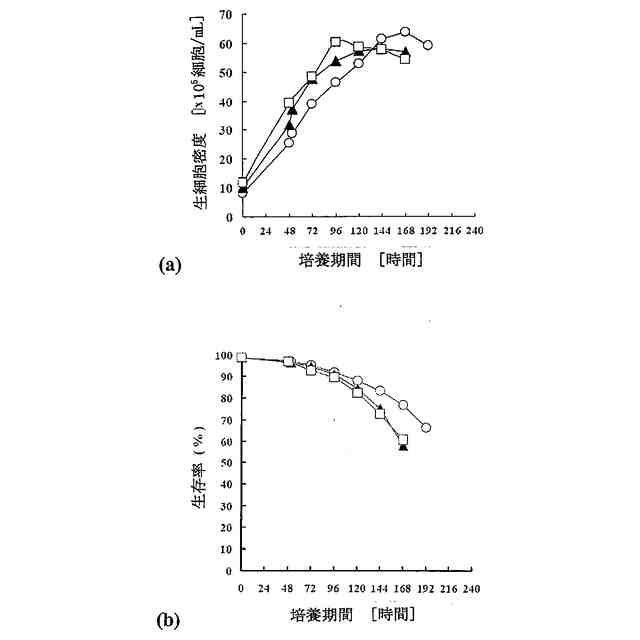
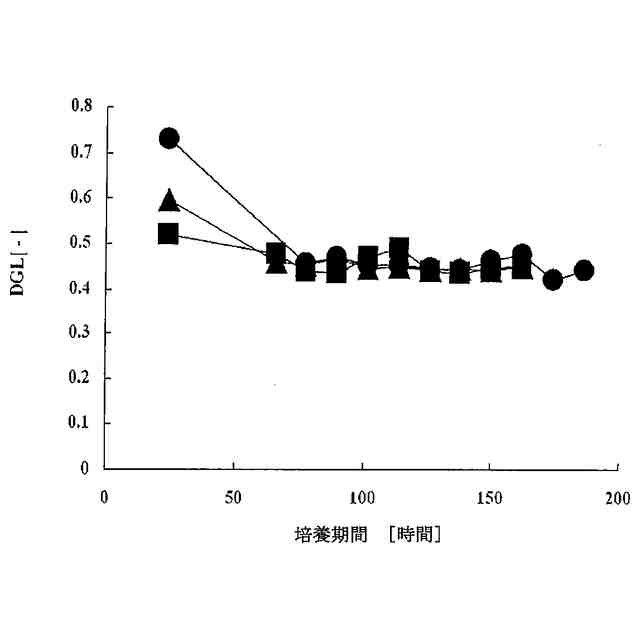
培養がフェドバッチ培養であることを特徴とする、前記請求項のいずれか1項に記載の方法。
【請求項7】
培養がフェドバッチ培養であり、その際、流加を培養2日目または3日目に開始することを特徴とする、請求項6に記載の方法。
【請求項8】
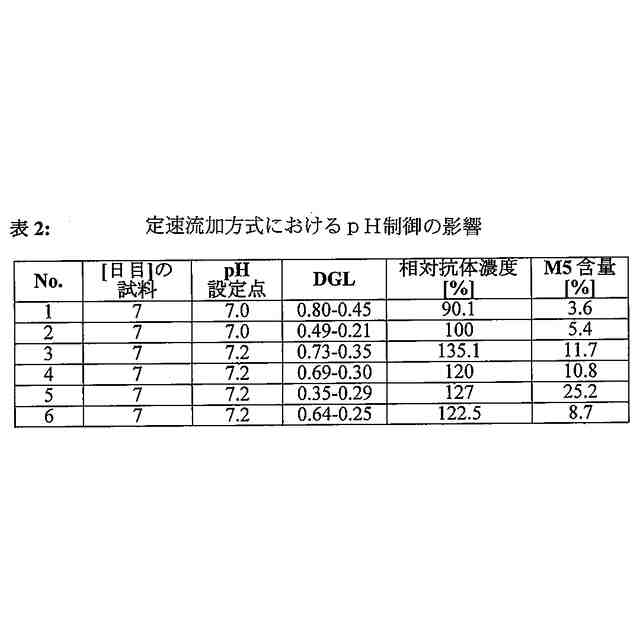
培養をpH6.5から7.5までのpH値で行なうことを特徴とする、前記請求項のいずれか1項に記載の方法。
【請求項9】
培養をpH6.9から7.3までのpH値で行なうことを特徴とする、請求項8に記載の方法。
【請求項10】
培養をpH6.95からpH7.05までのpH値、またはpH7.15からpH7.25までのpH値で行なうことを特徴とする、請求項9に記載の方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本明細書には、細胞における免疫グロブリン調製の分野の方法であって、調製される免疫グロブリンのグリコシル化パターンを培養条件に基づいて改変できる方法を記述する。
続きを表示(約 5,000 文字)
【背景技術】
【0002】
近年、免疫グロブリンの生産は常に増加しており、近い将来に免疫グロブリンは各種疾患の処置に利用できる最大グループの療法薬となるであろうと思われる。免疫グロブリンの影響はそれらの特異性から生じる;それには、それらの特異的な標的認識および結合の機能、ならびに抗原/Fc受容体の結合と同時または結合後の特異的作用の活性化が含まれる。
【0003】
特異的な標的認識および結合は、免疫グロブリンの可変部により仲介される。作用が生じる他の免疫グロブリン分子部分は、翻訳後修飾、たとえばグリコシル化パターンである。翻訳後修飾は実際に、免疫グロブリンの有効性、安定性、免疫原力潜在性、結合などに対して影響をもつ。それに関連して、補体依存性細胞傷害性(complement-dependent cytotoxicity)(CDC)、抗体依存性細胞性細胞傷害性(antibody-dependent cellular cytotoxicity)(ADCC)、およびアポトーシス誘導に対処しなければならない。
【0004】
免疫グロブリンのグリコシル化パターン、すなわち結合した糖構造体の糖の組成および数が生物学的特性に強い影響をもつことが報告された(たとえば、Jefferis, R., Biotechnol. Prog. 21 (2005) 11-16を参照)。哺乳動物細胞により産生される免疫グロブリンは
2~3質量%の炭水化物を含有する(Taniguchi, T., et al., Biochem. 24 (1985) 5551-5557)。これは、たとえばクラスGの免疫グロブリン(IgG)において、マウス由来の
IgGにおける2.3個のオリゴ糖残基(Mizuochi, T., et al., Arch. Biochem. Biophys. 257 (1987) 387-394)、およびヒト由来のIgGにおける2.8個のオリゴ糖残基と同等であり(Parekh, R.B., et al., Nature 316 (1985) 452-457)、これらのうち一般に2
個はFc領域、残りは可変部に位置する(Saba, J.A., et al., Anal. Biochem. 305 (2002) 16-31)。
【0005】
クラスGの免疫グロブリンのFc領域において、オリゴ糖残基はアミノ酸残基297、すなわちアスパラギン残基(Asn
297
と表記される)にN-グリコシル化により導入される可能性がある。Youingsらは、ポリクローナルIgG分子の15%~20%でFc
領域にさらに他のN-グリコシル化部位が存在することを示した(Youings, A., et al., Biochem. J., 314 (1996) 621-630;たとえば、Endo, T., et al., Mol. Immunol. 32 (1995) 931-940も参照)。不均一な、すなわち非対称的なオリゴ糖プロセシングのため、異なるグリコシル化パターンをもつ多数のイソ形免疫グロブリンが存在する(Patel, T.P., et
al., Biochem. J. 285 (1992) 839-845; Ip, C.C., et al., Arch. Biochem. Biophys. 308 (1994) 387-399; Lund, J., et al., Mol. Immunol. 30 (1993) 741-748)。同時に、オリゴ糖の構造および分布は高度に再現性があり(すなわち、非ランダム)、かつ部位特異的である(Dwek, R.A., et al., J. Anat. 187 (1995) 279-292)。
【0006】
免疫グロブリンの幾つかの特性はFc領域のグリコシル化に直接関連する(たとえば、Dwek, R.A., et al., J. Anat. 187 (1995) 279-292; Lund, J., et al., J. Immunol. 157 (1996) 4963-4969; Lund, J., FASEB J. 9 (1995) 115-119; Wright, A. and Morrison, S.L., J. Immunol. 160 (1998) 3393-3402を参照);たとえば、熱安定性および溶解性(West, C.M., Mol. Cell. Biochem. 72 (1986) 3-20)、抗原性(Turco, S.J., Arch. Biochem. Biophys. 205 (1980) 330-339)、免疫原性(Bradshaw, J.P., et al., Biochim. Biop
hys. Acta 847 (1985) 344-351; Feizi, T. and Childs, R.A., Biochem. J. 245 (1987)
1-11; Schauer, R., Adv. Exp. Med. Biol. 228 (1988) 47-72)、クリアランス速度/循環半減期(Ashwell, G. and Harford, J., Ann. Rev. Biochem. 51 (1982) 531-554; McFarlane, I.G., Clin. Sci. 64 (1983) 127-135; Baenziger, J.U., Am. J. Path. 121 (1985) 382-391; Chan, V.T. and Wolf, G., Biochem. J. 247 (1987) 53-62; Wright, A., et al., Glycobiology 10 (2000) 1347-1355; Rifai, A., et al., J. Exp. Med. 191 (2000) 2171-2182; Zukier, L.S., et al., Cancer Res. 58 (1998) 3905-3908)、ならびに特異的生物活性(Jefferis, R. and Lund, J., Antibody Engineering, ed. by Capra, J.D., Chem. Immunol. Basel, Karger, 65 (1997) 111-128)。
【0007】
グリコシル化パターンに影響を及ぼす要因が調べられた:たとえば、発酵培地中のウシ胎仔血清の存在(Gawlitzek, M., et al., J. Biotechnol. 42(2) (1995) 117-131)、緩衝作用条件(Muething, J., et al., Biotechnol. Bioeng. 83 (2003) 321-334)、溶存酸素
濃度(Saba, J.A., et al., Anal. Biochem. 305 (2002) 16-31; Kunkel, J.P., et al., J. Biotechnol. 62 (1998) 55-71; Lin, A.A., et al., Biotechnol. Bioeng. 42 (1993)
339-350)、オリゴ糖の位置およびコンホメーション、ならびに宿主細胞タイプおよび細
胞増殖状態(Hahn, T.J. and Goochee, C.F., J. Biol. Chem. 267 (1992) 23982-23987; Jenkins, N., et al., Nat. Biotechnol. 14 (1996) 975-981)、細胞のヌクレオチド-糖代謝(Hills, A.E., et al., Biotechnol. Bioeng. 75 (2001) 239-251)、栄養素制限(Gawlitzek, M., et al., Biotechnol. Bioeng. 46 (1995) 536-544; Hayter, P.M., et al.,
Biotechnol. Bioeng. 39 (1992) 327-335)、特にグルコース制限(Tachibana, H., et al., Cytotechnology 16 (1994) 151-157)、ならびに細胞外pH(Borys, M.C., et al., Bio/Technology 11 (1993) 720-724)。
【0008】
たとえばNS0骨髄腫細胞における免疫グロブリンの組換え発現により、オリゴマンノース構造体およびトランケート型オリゴ糖構造体の増加が観察された(Ip, C.C., et al.,
Arch. Biochem. Biophys. 308 (1994) 387-399; Robinson, D.K., et al., Biotechnol.
Bioeng. 44 (1994) 727-735)。グルコース飢餓条件下で、グリコシル化の変動、たとえ
ばより小さな前駆オリゴ糖の結合またはオリゴ糖部分の完全な不存在が、CHO細胞、ネズミ3T3細胞、ラット肝癌細胞、ラット腎細胞およびネズミ骨髄腫細胞において観察された(Rearick, J.I., et al., J. Biol. Chem. 256 (1981) 6255-6261; Davidson, S.K. and Hunt, L.A., J. Gen. Virol. 66 (1985) 1457-1468; Gershman, H. and Robbins, P.W., J. Biol. Chem. 256 (1981) 7774-7780; Baumann, H. and Jahreis, G.P., J. Biol.
Chem. 258 (1983) 3942-3949; Strube, K.-H., et al., J. Biol. Chem. 263 (1988) 3762-3771; Stark, N.J. and Heath, E.C., Arch. Biochem. Biophys. 192 (1979) 599-609)。低グルタミン/グルコース濃度に基づく方法がWong, D.C.F., et al., Biotechnol. Bioeng. 89 (2005) 164-177により報告された。
【0009】
日本国特許出願JP 62-258252には哺乳動物細胞の潅流培養が記述され、一方、US 5,443,968にはタンパク質分泌細胞のフェドバッチ(fed-batch)培養法(流加培養法)が報告
されている。WO 98/41611には、低い乳酸産生を特徴とする代謝状態に細胞を適応させる
のに有効な細胞培養法が報告されている。物質を産生させるために細胞を培養する方法がWO 2004/048556に報告されている。Elbein, A.D., Ann. Rev. Biochem. 56 (1987) 497-534には、グルコースの不存在下でインキュベートした哺乳動物細胞はマンノース-9含有構造体の代わりにマンノース-5含有構造体をタンパク質へ伝達することが報告されている。pCO2の依存性は、グルコース制限に際してCHO細胞の増殖、代謝およびIgG産生に影響を及ぼすことがTakuma, S., et al. in Biotechnol. Bioeng. 97 (2007) 1479-1488により報告されている。
【先行技術文献】
【特許文献】
【0010】
JP 62-258252
US 5,443,968
WO 98/41611
WO 2004/048556
【非特許文献】
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
合同酒精株式会社
麦汁の製造方法
10日前
マグネデザイン株式会社
磁気顕微鏡
2か月前
池田食研株式会社
RNAの合成方法
19日前
東洋紡株式会社
細菌からの核酸抽出法
25日前
株式会社ゴーフォトン
PCR方法
2か月前
熊本県
低褐変レタスとその作製方法
1か月前
東洋紡株式会社
ウイルスからの核酸抽出法
25日前
SMC株式会社
気体供給装置
2か月前
テルモ株式会社
液体除去器具
1か月前
国立大学法人山梨大学
受精胚の選別方法及び装置
1か月前
朝日酒造 株式会社
発泡性清酒の製造方法
22日前
鹿島建設株式会社
褐藻の冷凍保存方法
2か月前
セージ セラピューティクス, インコーポレイテッド
C7、C12、およびC16置換神経刺激性ステロイドおよびそれらの使用方法
16日前
株式会社テクノーブル
乳酸菌及び皮膚外用剤
2か月前
サッポロビール株式会社
アルコール飲料
2か月前
大和ハウス工業株式会社
藻類培養システム
18日前
株式会社SUPER BLOOM
スピリッツ
2か月前
テルモ株式会社
移植用デバイス
1か月前
新東工業株式会社
培養システム
2か月前
新東工業株式会社
培養システム
2か月前
株式会社ユーグレナ
観察用遠隔制御装置
1か月前
テルモ株式会社
積層体および移植片の輸送方法
1か月前
サッポロビール株式会社
茶風味アルコール飲料
2か月前
住友化学株式会社
細胞用基材
1か月前
日本臓器製薬株式会社
髄核細胞の培養方法
1か月前
住友化学株式会社
細胞用基材
1か月前
三井化学株式会社
作業用装置
1か月前
株式会社アステック
培養装置及び培養方法
1か月前
住友化学株式会社
細胞チップ
9日前
学校法人慈恵大学
培養皿用の定規
22日前
株式会社関電工
藻類培養装置および藻類培養方法
1か月前
TOA株式会社
化粧品の保存効力試験方法
1か月前
国立大学法人愛媛大学
ASC欠損非ヒト哺乳動物
1か月前
国立研究開発法人産業技術総合研究所
細胞観察方法及びその装置
1か月前
有限会社栄和商事
藻類の培養方法及び藻類培養装置
1か月前
宝酒造株式会社
蒸留酒類、及び蒸留酒類の製造方法
1か月前
続きを見る
他の特許を見る




 特許ウォッチ
特許ウォッチ