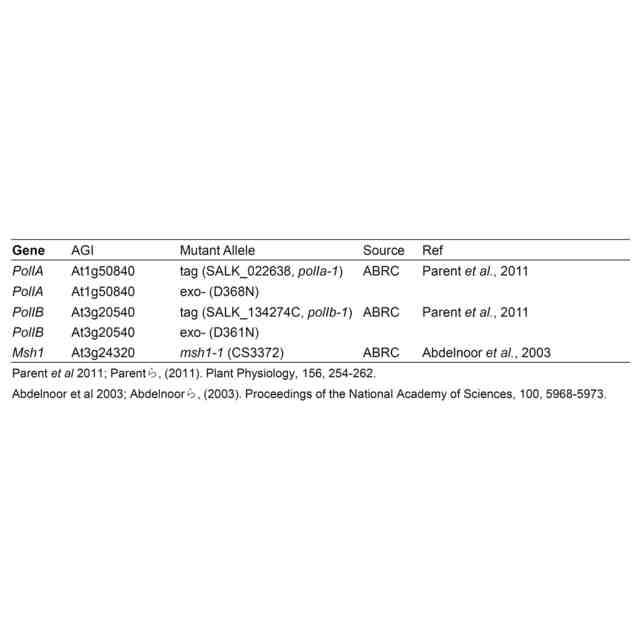
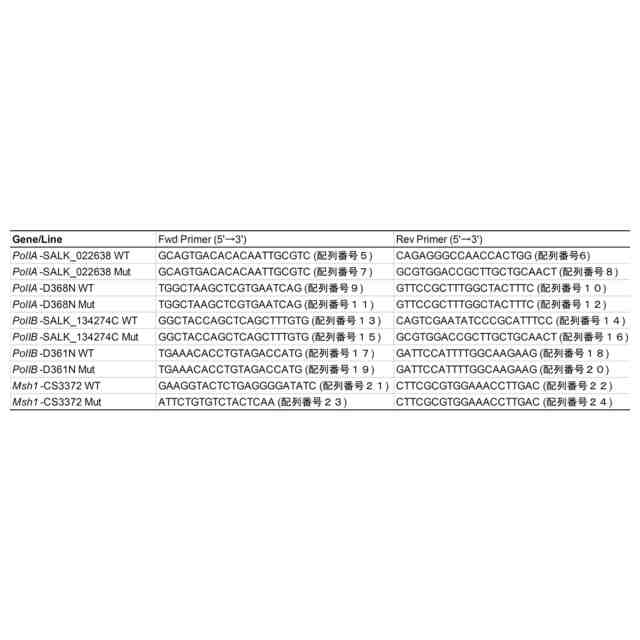
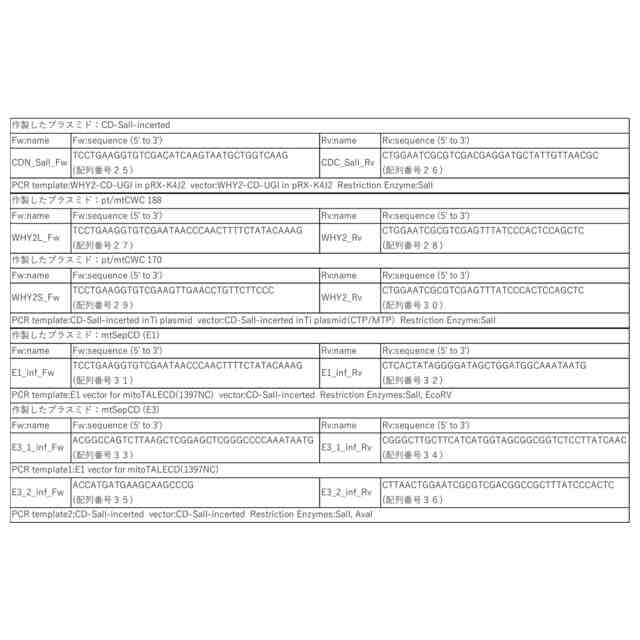
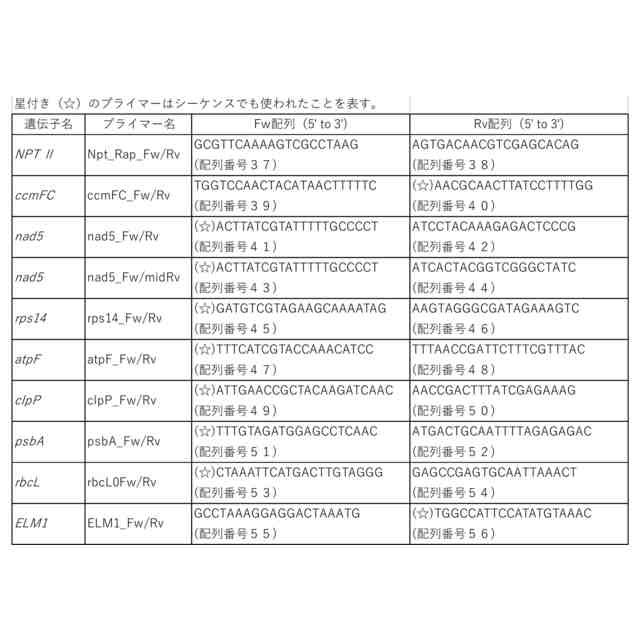
特許請求の範囲【請求項1】 ゲノムDNAにランダムな変異を導入する方法であって、以下の(a)および/または(b)の工程を含む方法。 (a)DNAポリメラーゼのDNA複製エラー頻度を上昇させる工程、 (b)塩基置換酵素、または塩基置換酵素と配列非特異的DNA結合因子の複合体を、細胞内に導入する工程。 続きを表示(約 950 文字)【請求項2】 ゲノムDNAにランダムな変異が導入された細胞を作製する方法であって、以下の(a)および/または(b)の工程を含む方法; (a)DNAポリメラーゼのDNA複製エラー頻度を上昇させる工程、 (b)塩基置換酵素、または塩基置換酵素と配列非特異的DNA結合因子の複合体を、細胞内に導入する工程。 【請求項3】 前記DNAポリメラーゼのエキソヌクレアーゼ活性を低下または喪失させることにより、DNAポリメラーゼのDNA複製エラー頻度を上昇させることを特徴とする、請求項1または2に記載の方法。 【請求項4】 前記ゲノムがオルガネラのゲノムである、請求項3に記載の方法。 【請求項5】 前記エキソヌクレアーゼ活性を低下または喪失させる方法が、オルガネラDNAポリメラーゼのExoII領域に存在するアスパラギン酸に変異を加えることである、請求項4に記載の方法。 【請求項6】 前記変異がアスパラギン酸からアスパラギンへの置換である、請求項5に記載の方法。 【請求項7】 前記DNAポリメラーゼのポリメラーゼドメインに変異を導入することにより、DNAポリメラーゼのDNA複製エラー頻度を上昇させることを特徴とする、請求項1または2に記載の方法。 【請求項8】 前記塩基置換酵素がシチジンデアミナーゼである、請求項1または2に記載の方法。 【請求項9】 前記シチジンデアミナーゼが、以下の(a)または(b)に記載のいずれかのタンパク質である、請求項8に記載の方法; (a)配列番号1で表されるアミノ酸配列からなるタンパク質、 (b)配列番号1で表されるアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列からなり、かつ、シチジンデアミナーゼ活性を有するタンパク質。 【請求項10】 前記シチジンデアミナーゼと前記配列非特異的DNA結合因子との複合体が、シチジンデアミナーゼのN末端の一部分とそれ以外の部分との間に当該配列非特異的DNA結合因子が融合されているものである、請求項9に記載の方法。 (【請求項11】以降は省略されています) 発明の詳細な説明【技術分野】 【0001】 本発明は、ゲノムDNAへランダムに変異を導入する方法に関する。 続きを表示(約 3,400 文字)【背景技術】 【0002】 植物や動物の育種法の1つとして、ランダム変異原処理を行った突然変異育種法がある。この方法は、何らかの方法でゲノム(核ゲノムおよびオルガネラゲノム)に損傷を与えることで、ゲノムの塩基置換や塩基の挿入欠失を引き起こし、生じた変異集団の中から望ましい形質を示す個体を選抜する方法で、いわゆる順遺伝学的なアプローチとして知られている。植物ゲノムへのランダム変異の導入は、初期の頃は、X線を用いて実施されていたが、その後、メタンスルホン酸エチル(Ethyl methane sulfonate:EMS)やN-ニトロソ-N-メチル尿素(N-Methyl-N-nitrosourea:NMU)などの化学変異原処理、γ線をはじめとする放射線による処理などが多く用いられるようになってきた。 【0003】 核にコードされた遺伝子やタンパク質に関する知見が蓄積されると、重要なタンパク質の機能を改善する「指向性進化」のために、特定の遺伝子領域に絞ったランダム変異(focused mutagenesis)を導入するerror prone PCRやDNAシャッフリングが行われるようになった。また、CRISPR/Cas9やBase editorに代表される「ゲノム編集」ツールにより、ゲノムDNAの一部だけを改変し、求める変異体を効率的に得ることが出来るようになった。ゲノム編集ツールであるCRISPR/Cas9やBase editorは指向性進化法のためのツールとしても応用され始めており、突然変異誘発の大きな流れはfocused mutagenesisに向いている。 【0004】 核ゲノムの遺伝子で逆遺伝学や指向性進化が盛んにおこなわれる一方、近年細胞質に存在するミトコンドリア(mt)ゲノムにおけるmitoTALENによる標的遺伝子の破壊(非特許文献1、非特許文献2)や、色素体(pt)ゲノムとmtゲノムの両方における標的一塩基置換(非特許文献3、非特許文献4、特許文献1)、そしてmitoTALENを応用したmtゲノム内のfocused mutagenesis(非特許文献5)が達成された。 【0005】 しかし、オルガネラゲノムには、遺伝子自体の機能やアミノ酸配列と機能の関係性、未知のORF(open reading frame)、遺伝子のシス配列・遺伝子間配列の役割など、多くの未解明点が残されている。開発された上記のオルガネラゲノムの直接改変法はそれらの未解明点を調べる有力な手段であるが、両オルガネラゲノム上のすべての遺伝子や遺伝子間領域を標的として改変を行うことは現実的ではない。そのため、知見の不足している段階で特に有力なゲノムワイドなランダム変異導入技術は、オルガネラゲノムに関する研究やそこから翻訳されるタンパク質の機能改善に重要な役割を果たすと言える。 【0006】 ptゲノムに関して、ランダム変異導入のための方法はいくつか報告されており、化学変異原処理による直接変異導入と核コードの遺伝子変異による副次的変異導入の二つに大別される。前者の例として、1960年台後半よりNMUを使った複数の高等植物へのptゲノム改変が行われ、以降標準的に用いられるようになった(非特許文献6)。また、核ゲノムへのランダム変異導入剤として一般的なEMSによっても、タバコ(Nicotiana tabacum)のルビスコ大サブユニット遺伝子(rbcL)点変異系統の創出(非特許文献7)や、赤トウガラシ(Capsicum annuum L.)の葉緑体コード遺伝子改変による抗生物質耐性の獲得が行われている(非特許文献8)。後者は主にオルガネラゲノム修復関連の核にコードされた遺伝子を変異させることによる。当初chloroplast mutator(chm)として単離されたMutS Homolog 1(msh1)変異が有名であるが、この遺伝子の変異はptゲノム、mtゲノムの両方に影響することが後に明らかとなった(非特許文献9)。また、why1why3二重変異体はptゲノム特異的な変異を引き起こすとされるが、それらは短い相同配列間の組換えを用いた誤修復による構造変化であると報告された(非特許文献10)。このように複数の報告がされている中で、ptゲノム特異的にランダムに点変異を導入する効率的な手法は確立していない。 【0007】 また、mtゲノムに関するランダム変異導入はより報告が少なく、二品種のタバコ細胞を融合させて新規のmtゲノム構造を得た例(非特許文献11)や、RNAiノックダウンのmsh1変異体における構造変化が引き起こすCMS(cytoplasmic male sterility)個体の創出(非特許文献12)、T-DNA挿入によるmsh1ホモ変異体とreca3ホモ変異体における構造変化(非特許文献13)、および複数のT-DNA挿入msh1ホモ変異体における野生型の100-1000倍の挿入欠失、10倍の塩基置換、構造変化(非特許文献9)などが挙げられるが、mtゲノムへ特異的に点変異を導入することは困難である。 【0008】 以上のように、オルガネラゲノムに対して、多様な突然変異変化を導入する技術は、未だに確立しておらず、さらなる改善された技術の開発が必要とされている。また、核ゲノムについても、新たな効率的な突然変異の導入法が望まれている。 【先行技術文献】 【特許文献】 【0009】 WO2022/158561A1 【非特許文献】 【0010】 Kazamaら, (2019) Nature Plants, 5, 722-730. Arimuraら, (2020) The Plant Journal: For Cell and Molecular Biology, 104, 1459-1471. Nakazatoら, (2021) Nature Plants, 7, 906-913. Nakazatoら, (2022) Proceedings of the National Academy of Sciences of the United States of America, 119, e2121177119. Fornerら, (2022) Nature Plants, 8, 245-256. Fluhrら, (1985) Proceedings of the National Academy of Sciences of the United States of America, 82, 1485-1489. Shikanaiら, (1996) Plant Molecular Biology, 31, 399-403. Subhashら, (1997) In Vitro Cellular & Developmental Biology-Plant, 33, 285-287. Wuら, (2020) Proceedings of the National Academy of Sciences of the United States of America, 117, 16448-16455. Marechalら, (2009) Proceedings of the National Academy of Sciences of the United States of America, 106, 14693-14698. Belliardら, (1979) Nature, 281, 401-403. Sandhuら, (2007) Proceedings of the National Academy of Sciences of the United States of America, 104, 1766-1770. Shedgeら, (2007) The Plant Cell, 19, 1251-1264. 【発明の概要】 【発明が解決しようとする課題】 (【0011】以降は省略されています) この特許をJ-PlatPatで参照する



 特許ウォッチ
特許ウォッチ