TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025078673
公報種別
公開特許公報(A)
公開日
2025-05-20
出願番号
2025030446,2022557169
出願日
2025-02-27,2021-03-19
発明の名称
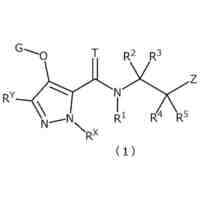
インダゾール系化合物及び関連使用方法
出願人
アルヴィナス・オペレーションズ・インコーポレイテッド
代理人
個人
,
個人
主分類
C07D
401/14 20060101AFI20250513BHJP(有機化学)
要約
【課題】LRRK2に関連する疾患及び障害、例えば、特発性PD、LRRK2変異関連PD(例えば、1つ以上のLRRK2活性化変異に関連するPD)、原発性タウオパチー(例えば、核上性麻痺(PSP)または皮質基底核変性症(CBD))、レビー小体型認知症、クローン病、ハンセン病(例えば、1型炎症性反応を伴うハンセン病)、及び/または神経炎症への有効な治療に対する継続的な必要性が当該技術分野において存在する。
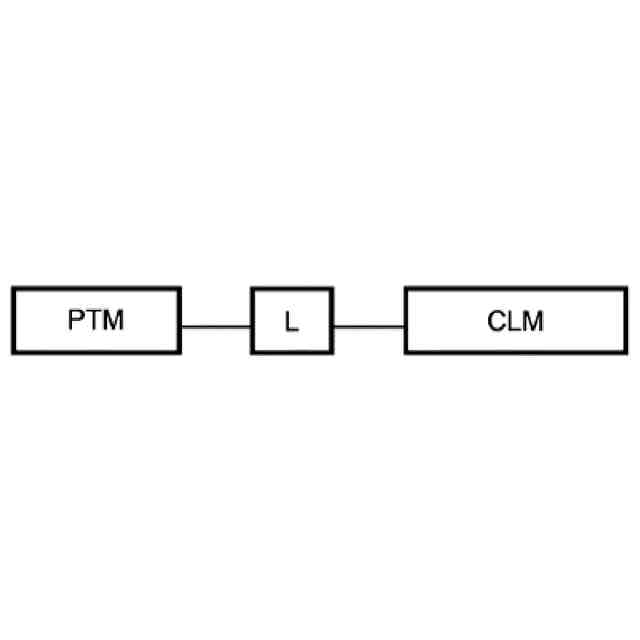
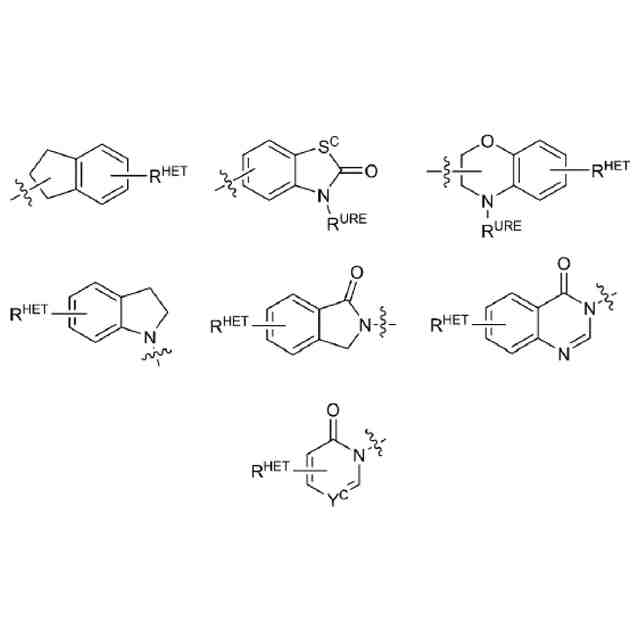
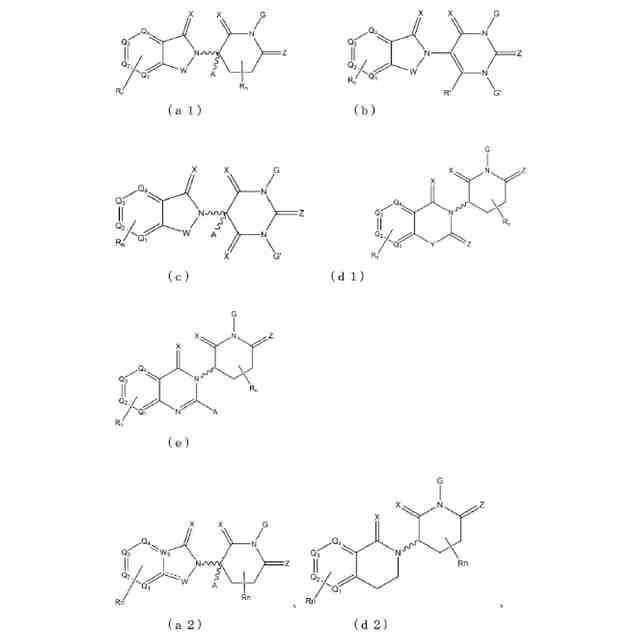
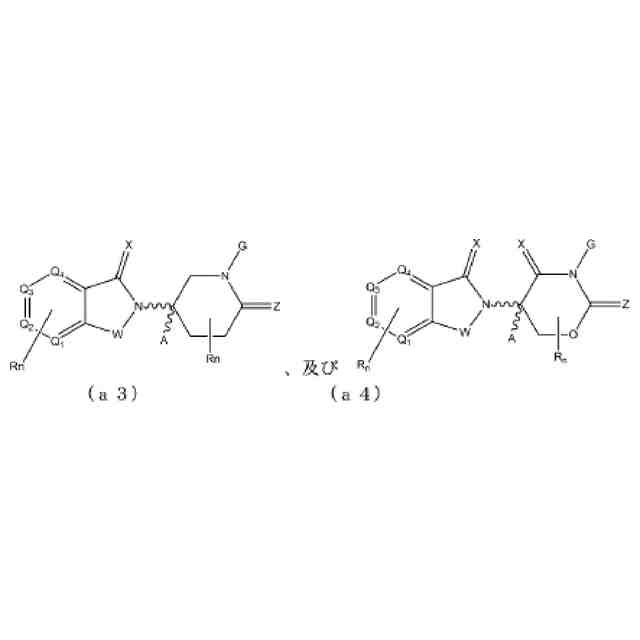
【解決手段】ロイシンリッチリピートキナーゼ2(LRRK2)のモジュレーターとして有用である二機能性化合物が本明細書に記載される。特に、本開示のヘテロ二機能性化合物は、一端にセレブロンE3ユビキチンリガーゼに結合する部分を含有し、他端にLRRK2に結合する部分を含有することから、標的タンパク質がユビキチンリガーゼに近接して配置され、標的タンパク質の分解(及び阻害)がもたらされる。本開示のヘテロ二機能性化合物は、標的タンパク質の分解/阻害に関連する幅広い薬理学的活性を呈する。標的タンパク質の異常な調節に起因する疾患または障害が本開示の化合物及び組成物によって治療または予防される。
【選択図】なし
特許請求の範囲
【請求項1】
本明細書に記載された発明。
発明の詳細な説明
【技術分野】
【0001】
関連出願の相互参照
本開示は、2020年3月21日に出願されたインダゾール系化合物及び関連使用方法というタイトルの米国特許仮出願第62/992,952号の利益及び優先権を主張するものであり、全ての目的のために、その全体が参照により本明細書に援用される。
続きを表示(約 9,600 文字)
【0002】
参照による援用
引用される全ての参考文献は、米国特許出願公開第2015/0291562号として公開された2015年4月14日出願の米国特許出願第14/686,640号;及び米国特許出願公開第2016/0058872号として公開された2015年7月6日出願の米国特許出願第14/792,414号;及び米国特許出願公開第2018/0228907号として公開された2018年4月13日出願の米国特許出願第15/953,108号;及び2015年9月2日に出願された米国特許出願公開第2016/0009689A1号;及び2016年2月18日に出願された米国特許出願公開第2016/0200722A1号を含め、その全体が参照により本明細書に援用される。
本発明は、標的タンパク質結合部分及びE3ユビキチンリガーゼ結合部分を含むヘテロ二機能性化合物、ならびに関連使用方法を提供する。二機能性化合物は、ロイシンリッチリピートキナーゼ2(LRRK2)の標的化されたユビキチン化のモジュレーターとして有用であり、LRRK2は、次いで、分解及び/または阻害される。
【背景技術】
【0003】
ほとんどの低分子薬物は、酵素または受容体に、ぴったりとした明確に画定されたポケットで結合する。一方、タンパク質間相互作用は、接触表面が広く、浅い溝または平坦な界面が関与することから、小分子を使用して標的化することが難しいことで知られている。E3ユビキチンリガーゼ(ヒトでは数百種が知られている)は、ユビキチン化に対する基質特異性を与えることから、特定のタンパク質基質に対する特異性があるため、一般的なプロテアソーム阻害剤よりも魅力的な治療標的である。E3リガーゼのリガンドの開発は、一部には、タンパク質間相互作用を阻止しなければならないという事実により、困難であることが分かっている。しかしながら、近年の開発により、これらのリガーゼに結合する特異的なリガンドが提供されるようになった。例えば、最初の低分子E3リガーゼ阻害剤であるヌトリンの発見以来、E3リガーゼを標的とする更なる化合物が報告されている。
【0004】
セレブロンは、ヒトではCRBN遺伝子によってコードされているタンパク質である。CRBNのオルソログは、植物からヒトまで高度に保存されており、その生理学的な重要性が強く示唆される。セレブロンは、損傷DNA結合タンパク質1(DDB1)、カリン4A(CUL4A)、及びカリン調節因子1(ROC1)とともにE3ユビキチンリガーゼ複合体を形成する。この複合体は、多数の他のタンパク質をユビキチン化する。まだ完全に解明されていない機構を介したセレブロンによる標的タンパク質のユビキチン化は、線維芽細胞増殖因子8(FGF8)及び線維芽細胞増殖因子10(FGF10)のレベル上昇をもたらす。次いで、FGF8は、四肢及び耳胞形成などの多数の発生過程を調節する。最終結果として、このユビキチンリガーゼ複合体は、胚における四肢の成長に重要である。セレブロンが存在しない場合、DDB1は、DNA損傷結合タンパク質として機能するDDB2と複合体を形成する。
【0005】
米国特許出願公開第2015/0291562号及び同第2014/0356322号(参照により本明細書に援用される)に記載されるものなどの二機能性化合物は、ユビキチン化と、それに続くプロテアソーム分解経路での分解のために、内因性タンパク質をE3ユビキチンリガーゼに動員するように機能する。特に、上に引用される公開物には、二機能性またはタンパク質分解誘導キメラ(PROTAC(登録商標))タンパク質分解化合物が記載され、様々なポリペプチド及びタンパク質の標的ユビキチン化のモジュレーターとしての有用性が見出されており、次いで、二機能性化合物によって分解及び/または阻害される。
【0006】
ロイシンリッチリピートキナーゼ2(LRRK2)は、ロイシンリッチリピートキナーゼファミリーのメンバーであり、N末端のアルマジロドメイン、アンクリンリピート領域、ロイシンリッチリピート(LRR)ドメイン、タンデムRoco型GTPaseドメイン、DFG様モチーフ含有キナーゼドメイン、及びC末端のWD40ドメインを有する大きなマルチドメインタンパク質である。LRRK2タンパク質は、2527アミノ酸及び分子量280kDaである。LRRK2の触媒活性は、キナーゼドメイン及びGTPaseドメインに関連し、LRRK2は、その活性型ではヘテロ二量体である(Greggio E,et al.:The Parkinson disease-associated leucine-rich repeat kinase 2(LRRK2)is a dimer that undergoes intramolecular autophosphorylation.J Biol Chem 2008,283:16906-16914)。GTP結合は、キナーゼ活性に必須であり、GTP結合を阻害する変異により、LRRK2キナーゼの活性が損なわれることが示されている(Ito G,et al.:GTP binding is essential to the protein kinase activity of LRRK2,a causative gene product for familial Parkinson’s disease.Biochemistry 2007,46:1380-1388)。唯一確認されている生理学的基質(LRRK2自体以外)は、Rab8a及びRab10を含む低分子量のGタンパク質のサブセットであり、これらは、小胞輸送及びエンドソーム機能の調節、ならびに細胞骨格ネットワーク上での輸送に関与している(Steger M,et al.:Phosphoproteomics reveals that Parkinson’s disease kinase LRRK2 regulates a subset of Rab GTPases.Elife 2016,5.e12813)。LRRK2の発現レベルは、免疫細胞(好中球、単球及びB細胞)、肺及び腎臓で最も高く、脳では発現量が低く、黒質のドーパミン作動性ニューロンで発現している(West AB,et al.:Differential LRRK2 expression in the cortex,striatum,and substantia nigra in transgenic and nontransgenic rodents.J Comp Neurol 2014,522:2465-2480)。
【0007】
LRRK2には、いくつかの優性な機能獲得型の病原性及び特徴的な変異があり、GTP加水分解に影響するRocoドメイン(N1437H、R1441G/C/H、Y1699C)、またはキナーゼドメイン(G2019S及びI2020T)のいずれかに位置する。G2019Sは、安静時振戦、筋強剛、運動減少(運動緩慢)、及び姿勢保持障害を特徴とする進行性神経変性障害である、パーキンソン病(PD)に関連する最も一般的なLRRK2変異である。PDの組織学的な特徴には、黒質緻密部におけるドーパミン作動性ニューロンの神経変性、ならびにレビー小体と呼ばれる細胞内封入体及びアルファ-シヌクレインタンパク質の凝集体からなる神経突起が含まれる。G2019Sは、全PD患者の1~2%に関連し、in vitroで2倍のキナーゼ活性の増加を引き起こし(West AB,et al.:Parkinson’s diseaseassociated mutations in leucine-rich repeat kinase 2 augment kinase activity.Proc Natl Acad Sci U S A 2005,102:16842-16847)、Ser1292での自己リン酸化が4倍増加する(Sheng Z,et al.:Ser1292 autophosphorylation is an indicator of LRRK2 kinase activity and contributes to the cellular effects of PD mutations.Sci Transl Med 2012,4:164ra161)。G2019S及びI2020Tの変異は、全てのキナーゼに共通するDFGモチーフ(LRRK2の場合DYGI)内にあり、触媒活性を制御する。これらの変異は、不活性型コンフォメーションを崩すことで、触媒活性を高めると考えられる(Schmidt SH,et al.:The dynamic switch mechanism that leads to activation of LRRK2 is embedded in the DFGpsi motif in the kinase domain.Proc Natl Acad Sci USA 2019,116:14979-14988)。上記のパーキンソン病関連変異のいくつか(R1441C/G、Y1699C及びI2020T)は、LRRK2のSer910及びSer935でのリン酸化を抑制し、その結果、LRRK2の不活性型を表すと考えられるLRRK2と14-3-3タンパク質との会合が減少する(Nichols J,et al.:14-3-3 binding to LRRK2 is disrupted by multiple Parkinson’s disease associated mutations and regulates cytoplasmic localisation.Biochem J 2010,430:393-404)。
【0008】
更に、LRRK2は、LRRK2遺伝子に関連するPARK8と呼ばれる12番染色体の領域内の変異により、常染色体優性遺伝性PDに関連している(Funayama M,et al.:A new locus for Parkinson’s disease(PARK8)maps to chromosome 12p11.2-q13.1.Ann Neurol 2002,51:296-301;Zimprich A,et al.:Mutations in LRRK2 cause autosomal-dominant parkinsonism with pleomorphic pathology.Neuron 2004,44:601-607;Paisan-Ruiz C,et al.:Cloning of the gene containing mutations that cause PARK8-linked Parkinson’s disease.Neuron 2004,44:595-600)。LRRK2は、1978年に初めて常染色体優性遺伝性パーキンソン病との関連性が指摘され、日本のある家族で追跡された(Nukada H,et al.:[A big family of paralysis agitans(author’s transl)].Rinsho Shinkeigaku 1978,18:627-634)。最も一般的な病原性LRRK2変異(G2019S)は、家族性PD症例の4~8%及び孤発性PD症例の1~3%で生じている。加えて、G2019S変異は、選択した家系のPD患者で共通しており、北アフリカ系ベルベル人で30~40%及びユダヤ系患者で14%がこの変異を保有している。
【0009】
LRRK2キナーゼ阻害剤は、G2019SなどのLRRK2活性が増加する変異誘発性PD、及びLRRK2の活性が増加する特発性PDを治療する可能性が提案されている(Chen J,et al.:Leucine-rich repeat kinase 2 in Parkinson’s disease:updated from pathogenesis to potential therapeutic target.Eur Neurol 2018,79:256-265;Alessi DR,et al.:LRRK2 kinase in Parkinson’s disease.Science 2018,360:36-37;Di Maio R,et al.:LRRK2 activation in idiopathic Parkinson’s disease.Sci Transl Med 2018,10)。いくつかの治療薬は、臨床に進んでおり、例えば、下流標的のリン酸化に直接影響するLRRK2キナーゼ阻害剤や、CNSに直接注入し、LRRK2タンパク質の翻訳を遮断することによってLRRK2タンパク質レベルを減少させるオリゴヌクレオチド(ASO)がある。
【0010】
レビー小体は、PDの主要な組織学的特徴である。レビー小体は、主にアルファ-シヌクレイン凝集体で構成されており、この凝集を増加させるアルファ-シヌクレインの変異もまたPDの発生リスクを増加させる(Meade RM,et al.:Alpha-synuclein structure and Parkinson’s disease lessons and emerging principles.Mol Neurodegener 2019,14.29-29)。ASOによるLRRK2の枯渇(Zhao HT,et al.:LRRK2 antisense oligonucleotides ameliorate a-synuclein inclusion formation in a Parkinson’s disease mouse model.Molecular therapy.Nucleic acids 2017,8:508-519)及びゲノムレベルでのLRRK2の欠失は、PDのマウスモデルにおいて、アルファ-シヌクレインを介した病態を減少させることが示されている(Lin X,et al.:Leucine-rich repeat kinase 2 regulates the progression of neuropathology induced by Parkinson’s-disease-related mutant alpha-synuclein.Neuron 2009,64:807-827)。G2019SなどのLRRK2活性を増加させる変異は、ニューロン及びPDのマウスモデルにおいて、アルファ-シヌクレインの凝集を増加させる。この増加は、LRRK2キナーゼ阻害剤により逆転した(Volpicelli-Daley LA,et al.G2019S-LRRK2 Expression Augments α-Synuclein Sequestration into Inclusions in Neurons.J Neurosci.2016 Jul 13;36(28):7415-27.doi:10.1523/JNEUROSCI.3642-15.2016)。LRRK2のG2019S変異体形態は、CNSにおいてキナーゼ阻害剤による阻害に耐性があり、その疾患修飾作用を低下させる可能性があることを示唆する証拠がいくつかある(Kelly K,et al.The G2019S mutation in LRRK2 imparts resiliency to kinase inhibition.Exp Neurol.2018 Nov;309:1-13)。PDのほとんどの症例では、死後検査でレビー小体も認めるが、PD症例に関連するLRRK2 G2019S変異の多くにはレビー小体は存在しない(Kalia LV,et al.:Clinical correlations with Lewy body pathology in LRRK2-related Parkinson disease.JAMA neurol 2015,72:100-105)。PDの共通の特徴であるレビー小体に加え、タウ病理も死後におけるLRRK2変異保持者の主な特徴である(Henderson MX,et al.:Alzheimer’s disease tau is a prominent pathology in LRRK2 Parkinson’s disease.Acta Neuropathol Commun 2019,7.183-183)。ある研究では、LRRK2変異保持者の100%でタウ病理が観察されたことから、LRRK2と核上性麻痺(PSP)または皮質基底核変性症(CBD)などの原発性タウオパチーとの遺伝的因果関係はさほど強くないものの、PDとの関係において、PDとタウ病理を結びつける重要な標的としてLRRK2が強調されている(Ross OA,et al.(2006)Lrrk2 R1441 substitution and progressive supranuclear palsy.Neuropathol Appl Neurobiol 32(1):23-25;Sanchez-Contreras M,et al.(2017)Study of LRRK2 variation in tauopathy:progressive supranuclear palsy and corticobasal degeneration.Mov Disord 32(1):115-123)。PSP生存の遺伝的決定因子として、LRRK2遺伝子座に共通する変異が近年報告された(Jabbari E,et al..,Common variation at the LRRK2 locus is associated with survival in the primary tauopathy progressive supranuclear palsy.bioRxiv 2020.02.04.932335;doi:https://doi.org/10.1101/2020.02.04.932335)。発現量的形質遺伝子座(eQTL)解析によるPSPでのLRRK2の発現増加は、反応性ミクログリアが誘発する炎症性状態をもたらし得、それにより、ミスフォールドタウタンパク質の継続的な蓄積及び臨床上の疾患進行を駆動することが報告されている。LRRK2の機能性バリアントはまた、クローン病及びハンセン病の1型炎症性反応とも関連している(Hui KY,et al..Functional variants in the LRRK2 gene confer shared effects on risk for Crohn’s disease and Parkinson’s disease.Sci Transl Med.2018 Jan 10;10(423).pii:eaai7795.doi:10.1126/scitranslmed.aai7795;Fava et al.Pleiotropic effects for Parkin and LRRK2 in leprosy type-1 reactions and Parkinson’s disease.Proc Natl Acad Sci U S A.2019 Jul 30;116(31):15616-15624.doi:10.1073/pnas.1901805116.Epub 2019 Jul 15)。
LRRK2は、好中球、単球及びマクロファージの免疫系、ならびに脳のミクログリアで高度に発現しており、ミクログリアの活性化及びリソソーム分解過程の内因性調節のモジュレーターである(Ma et al.Genetic comorbidities in Parkinson’s disease.Hum Mol Genet.2014 Feb 1;23(3):831-41.doi:10.1093/hmg/ddt465.Epub 2013 Sep 20、これは、Schapansky et al The complex relationships between microglia,alpha-synuclein,and LRRK2 in Parkinson’s disease.Neuroscience.2015 Aug 27;302:74-88.doi:10.1016/j.neuroscience.2014.09.049.Epub 2014 Oct 2において概説されている)。PDの疾患過程またはLRRK2の変異を介してこれらの免疫細胞の活性化が長くなると、神経炎症が増加し、PD及び/またはタウ病理の発生リスクが高まり得る。抗TNF剤による治療は、炎症性腸障害のある患者において、PDの発症リスクを78%減少させることから(Peter I,et al.:Anti-tumor necrosis factor therapy and incidence of Parkinson disease among patients with inflammatory bowel disease.JAMA Neurol 2018)、炎症とPDの強い関連が示唆される。PDに加えて、LRRK2は、がん、ハンセン病、及びクローン病などの他の疾患にも関連している(Lewis PA,Manzoni C.LRRK2 and human disease:a complicated question or a question of complexes?(2012).Sci Signal.5(207),pe2)。
【先行技術文献】
【特許文献】
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
株式会社トクヤマ
グルコンアミド誘導体
1か月前
日本化薬株式会社
新規顕色剤及び記録材料
1か月前
日本化薬株式会社
新規顕色剤及び記録材料
1か月前
花王株式会社
ポリアミド系ポリマー
2か月前
日産化学株式会社
ピリジン化合物の製造方法
13日前
ダイキン工業株式会社
シラン化合物
1か月前
日本化薬株式会社
シアノ複素環化合物の製造方法
3か月前
個人
メタンガス生成装置およびメタンガス生成方法
3か月前
花王株式会社
新規ピリジニウム化合物
1か月前
株式会社トクヤマ
グルコンアミド誘導体の製造方法
1か月前
個人
メタンガス生成装置およびメタンガス生成方法
3か月前
株式会社トクヤマ
結晶形Iのリオシグアトの製造方法
2か月前
四国化成工業株式会社
エポキシ化合物およびその利用
7日前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
2か月前
国立大学法人京都大学
細胞質送達ペプチド
1か月前
ダイキン工業株式会社
分離方法
2か月前
石原産業株式会社
シアノイミダゾール系化合物の製造方法
2か月前
四国化成工業株式会社
テレフタル酸化合物およびその利用
7日前
四国化成工業株式会社
イソフタル酸化合物およびその利用
7日前
株式会社半導体エネルギー研究所
有機化合物、発光デバイス
2か月前
ダイキン工業株式会社
SF5含有シラン化合物
1か月前
小川香料株式会社
化合物及び香料組成物
1か月前
株式会社カネカ
プロピレンオキサイド(PO)製造システム
6日前
キヤノン株式会社
有機化合物及び有機発光素子
12日前
小川香料株式会社
化合物及び香料組成物
1か月前
石原産業株式会社
シクラニリプロールの製造中間体の製造方法
2か月前
株式会社アイティー技研
炭化水素の合成方法及び合成装置
2か月前
オリザ油化株式会社
新規化合物及びその用途
2か月前
株式会社トクヤマ
精製第4級アンモニウム化合物水溶液の製造方法
25日前
出光興産株式会社
プロピレンの製造方法
今日
キッコーマン株式会社
ナノポアタンパク質
2か月前
国立大学法人九州大学
重水素化化合物の製造方法
2か月前
国立大学法人 東京大学
アシルヒドラゾン誘導体
3か月前
株式会社レゾナック
C2化合物の製造方法
2か月前
国立医薬品食品衛生研究所長
新規ペプチド
1か月前
マナック株式会社
2-ナフタレン誘導体の製造方法
3か月前
続きを見る
他の特許を見る


 特許ウォッチ
特許ウォッチ