TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
公開番号
2024133660
公報種別
公開特許公報(A)
公開日
2024-10-02
出願番号
2024109766,2022090304
出願日
2024-07-08,2016-09-08
発明の名称
標的化したDNA配列の核酸塩基を特異的に変換するゲノム配列の改変方法及びそれに用いる分子複合体
出願人
国立大学法人神戸大学
代理人
個人
,
個人
,
個人
主分類
C12N
15/09 20060101AFI20240925BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】宿主細胞内の二本鎖DNAの標的化された部位を改変する方法を提供すること。
【解決手段】本発明は、宿主細胞内の二本鎖DNAの標的化された部位を改変する方法であって、(a) 選択された二本鎖DNA中の標的ヌクレオチド配列の標的鎖に相補的な配列を含むcrRNAをコードするDNAと、(b)Cascadeを構成するタンパク質群と核酸塩基変換酵素とをコードするDNAであって、該核酸塩基変換酵素が該タンパク質群のいずれかのタンパク質と複合体を形成し得る形態で構成されたDNAとを、該宿主細胞に導入することにより、該標的化された部位において該二本鎖DNA鎖を切断することなく、該標的化された部位の1以上のヌクレオチドを他の1以上のヌクレオチドに変換する又は欠失させる、あるいは該標的化された部位に1以上のヌクレオチドを挿入する工程を含む、方法を提供する。
【選択図】図3
特許請求の範囲
【請求項1】
図面に記載の発明。
発明の詳細な説明
【技術分野】
【0001】
本発明は、DNAの二重鎖切断を伴わず、また外来DNA断片の挿入を行わずにゲノムの特定領域内の核酸塩基の改変を可能とする、ゲノム配列の改変方法及びそれに用いる、核酸配列認識モジュールと核酸塩基変換酵素との複合体に関する。
続きを表示(約 3,700 文字)
【背景技術】
【0002】
近年、様々な生物種において目的の遺伝子・ゲノム領域を改変する技術として、ゲノム編集が注目されている。従来、ゲノム編集の手法としては、配列非依存的なDNA切断能を有する分子と配列認識能を有する分子とを組み合わせた人工ヌクレアーゼを利用する方法が提案されている(非特許文献1)。
例えば、ジンクフィンガーDNA結合ドメインと非特異的なDNA切断ドメインとを連結した、ジンクフィンガーヌクレアーゼ(ZFN)を用い、宿主の植物細胞または昆虫細胞にDNA中の標的遺伝子座において組換えを行う方法(特許文献1)、植物病原菌キサントモナス属が有するDNA結合モジュールである転写活性化因子様(TAL)エフェクターと、DNAエンドヌクレアーゼとを連結したTALENを用いて、特定のヌクレオチド配列内又はそれに隣接する部位で、標的遺伝子を切断・修飾する方法(特許文献2)、あるいは、真正細菌や古細菌が持つ獲得免疫システムで機能するDNA配列CRISPR(ClusteredRegularly interspaced short palindromic repeats)と、CRISPRとともに重要な働きを持つヌクレアーゼCas(CRISPR-associated)タンパク質ファミリーとを組み合わせたCRISPR-Cas9システムを利用する方法(特許文献3)などが報告されている。さらには、35個のアミノ酸からなり1個の核酸塩基を認識するPPRモチーフの連続によって、特定のヌクレオチド配列を認識するように構成されたPPRタンパク質と、ヌクレアーゼとを連結した人工ヌクレアーゼを用い、該特定配列の近傍で標的遺伝子を切断する方法(特許文献4)も報告されている。
【0003】
これまで提案されてきたゲノム編集技術は、基本的にDNA二重鎖切断(double-stranded DNA breaks : DSB)を前提としているが、想定外のゲノム改変を伴うため、強い細胞毒性や染色体の転位などの副作用があり、遺伝子治療における信頼性を損なったり、ヌクレオチド改変による生存細胞数が極めて少なかったり、霊長類卵細胞や単細胞微生物では遺伝子改変自体が困難であるといった共通の課題があった。
【0004】
DSBを伴わないゲノム編集技術として、ジンクフィンガー(ZF)モチーフ等の配列認識能を有する分子と、核酸塩基のアミノ基をカルボニル基に変換するデアミナーゼとを組み合わせた人工酵素が提案されてはいるが(特許文献5)、実験的証拠は一切なく、変異導入効率はおろか、そもそも遺伝子改変が可能であるかすら明らかではない。実際、同じくゲノム切断を伴わない系として、DNA鎖上の脱塩基反応を触媒するDNAグリコシラーゼを用いた方法では、DNA修復機構を弱めた変異酵母を宿主に用いた場合ですら、変異導入効率は極めて低く(非特許文献2)、遺伝子治療や有用生物の分子育種等への実用化には程遠いのが現状である。
【先行技術文献】
【特許文献】
【0005】
特許第4968498号公報
特表2013-513389号公報
特表2010-519929号公報
特開2013-128413号公報
米国特許出願公開第2011/0104787号明細書
【非特許文献】
【0006】
Kelvin M Esvelt, Harris H Wang (2013) Genome-scale engineeri ng forsystems and synthetic biology, Molecular Systems Biology 9: 641
Prashant Mali, Kevin M Esvelt, George M Church (2013) Nuclei c AcidsRes. 41: e99
【発明の概要】
【発明が解決しようとする課題】
【0007】
従って、本発明の目的は、DSBないし外来DNA断片の挿入を伴わずに、遺伝子の特定配列
の核酸塩基を改変する、新規なゲノム編集の手法、並びにそのための核酸配列認識モジュール及び核酸塩基変換酵素の複合体を提供することである。
【課題を解決するための手段】
【0008】
本発明者らは既に、二本鎖DNAのいずれか一方もしくは両方の鎖の切断能を失活した変異Cas9を有するType II CRISPR-Casシステムを核酸配列認識モジュールとして用い、核酸塩基変換酵素としてデアミナーゼを用いることにより、DSBを伴うことなく、特定のDNA配列を含む領域においてゲノム配列を効率よく改変することに成功している(WO2015/133554)。しかし、Casタンパク質はprotospaceradjacent motif(PAM)と呼ばれるDNA鎖上の配列を認識して結合するため、変異導入部位はPAMの存否により制限される。現在、ゲノム編集に頻用されているTypeII CRISPR-Casシステムのストレプトコッカス・ピオゲネス(Streptococcus pyogenes)由来のCas9(SpCas9)は、PAMとしてNGG(Nは任意の塩基)を認識する。また、TypeII CRISPRにおいて標的とされるヌクレオチド配列(即ち、ガイドRNAに相補的なDNA鎖上のヌクレオチド配列)はPAMの5’上流18~25ヌクレオチドの長さであり、その長さにかかわらず、標的ヌクレオチド配列の5’端から2~5ヌクレオチドの位置で塩基変換が起こる頻度が最も高いことが、本発明者らの研究により明らかになっている。
【0009】
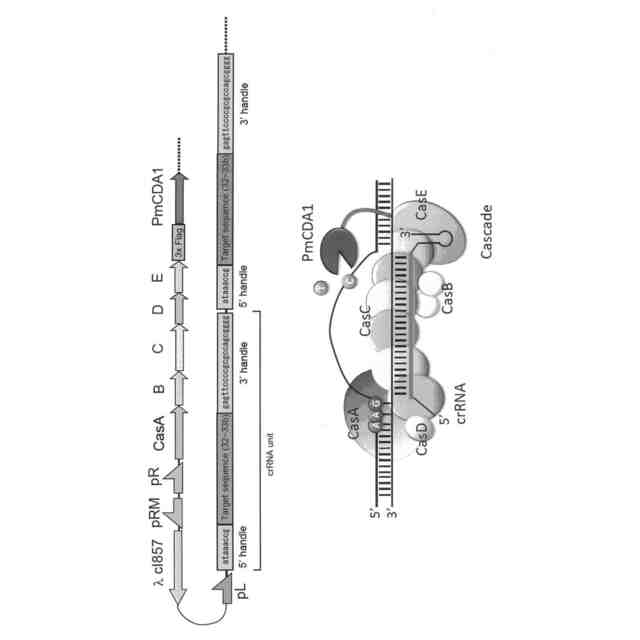
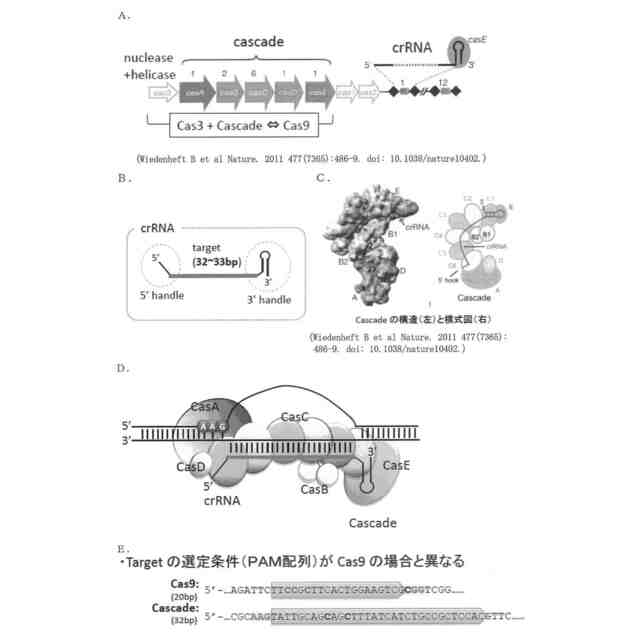
一方、大腸菌などが有するType I-E CRISPR-Casシステムでは、32~33ヌクレオチド長の標的ヌクレオチド配列の5’側に隣接するATG、AAG、AGG又はGAGを、PAM配列として認識することが知られており(図1参照;図1D及びEでは、PAM配列がAAGの場合を例示している)、他のTypeI CRISPR-Casシステムもそれぞれ特異的な2又は3塩基のPAM配列を認識する。従って、Type I CRISPR-Casシステムを核酸配列認識モジュールとして用いることができれば、Cas9ではPAM配列や変異導入位置の制約により変異導入が困難な部位での塩基変換が可能となる。
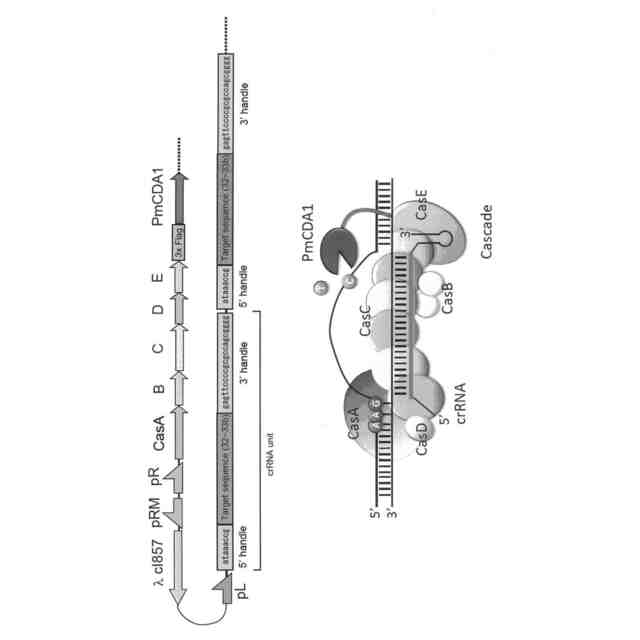
しかしながら、Type II CRISPR-Casシステムでは、標的ヌクレオチド配列に相補的なcrRNAとCas9をリクルートするためのtrans-actingcrRNA(tracrRNA)とのキメラRNAと、Cas9とのみから核酸配列認識モジュールを構成することができたが、例えば、大腸菌が有するType I-ECRISPR-Casシステムでは、標的ヌクレオチド配列とPAM配列の認識を、CRISPR-associated complex for antiviraldefence(Cascade)と呼ばれる、crRNAを提示した5種類のCasタンパク質(CasA、CasB、CasC、CasD及びCasE)とからなる複雑なリボヌクレオプロテイン複合体が担っており、これにヌクレアーゼ及びヘリカーゼ活性を有するCas3が加わって、TypeII CRISPR-CasシステムにおけるCas9と同様のDNA認識・切断機能を発揮している(図1参照)。このような複雑な構成のため、Type ICRISPR-Casシステムは、人工ヌクレアーゼとしてのゲノム編集技術にさえも、未だほとんど利用されていない。
【0010】
本発明者らは、大腸菌の必須遺伝子であるrpoB遺伝子の標的ヌクレオチド配列の標的鎖に相補的な配列の両端に、Cascadeに正しく提示されるのに必要な5’handle及び3’handleを連結したゲノム特異的CRISPR-RNA(crRNA)をコードするDNAを作製し、他方で、大腸菌のCasオペロンから、Cascadeの構成に必要なcasA~casE遺伝子群を単離し、これにデアミナーゼ遺伝子を連結したDNAを作製して、これらのDNAを、改変しようとする遺伝子を含む宿主大腸菌に導入した。その結果、目的遺伝子の標的ヌクレオチド配列及びその近傍に、ゲノムDNAの切断を伴うことなく変異を導入することに成功した。また、Cas9を用いた場合と異なり、標的ヌクレオチド配列の長さを32ヌクレオチドとすると、PAM配列から32~44塩基下流のシトシンが主に編集されることがわかった。
本発明者らは、これらの知見に基づいてさらに研究を重ねた結果、本発明を完成するに至った。
(【0011】以降は省略されています)
特許ウォッチbot のツイートを見る
この特許をJ-PlatPatで参照する
関連特許
国立大学法人神戸大学
細胞用の凍結保護剤
3か月前
国立大学法人神戸大学
糖鎖を有する重合体
2か月前
国立大学法人神戸大学
高効率な酵母遺伝子系
23日前
株式会社カネカ
結晶化促進剤
22日前
株式会社カネカ
耐衝撃性改良剤
2か月前
株式会社カネカ
海洋生分解促進剤
3か月前
国立大学法人神戸大学
中間バンドを用いた熱放射発電素子
3か月前
国立大学法人神戸大学
埋設管の埋設構造及び埋設管の埋設工法
3か月前
花王株式会社
3-ヒドロキシ安息香酸類の製造方法
2か月前
国立大学法人神戸大学
単相ダイレクトAC-ACコンバータとその電力制御方法
1か月前
国立大学法人神戸大学
抗SARS-CoV-2抗体、及びそれを含む検査キット
3か月前
国立大学法人 東京大学
血糖制御能力の推定方法及び装置
4か月前
住友ベークライト株式会社
冷凍生肉用真空包装体の製造方法
1か月前
JFEスチール株式会社
検査装置、検査システム、及び検査方法
3か月前
JFEスチール株式会社
検査装置、検査システム、及び検査方法
3か月前
株式会社ファーマフーズ
SARS-CoV-2変異株特異的認識抗体
2か月前
ENEOS株式会社
クレゾール又はベンズアルデヒドの製造方法、及び、触媒
3か月前
国立大学法人神戸大学
反共振を加えた多重共振形LLC共振タンク及びそれを用いたDC-DCコンバータ
3か月前
国立大学法人神戸大学
細胞の有する二本鎖DNAの標的部位を改変する方法
13日前
日本電信電話株式会社
学習装置、シミュレーション装置、学習方法、シミュレーション方法、及びプログラム
2か月前
国立大学法人神戸大学
潰瘍性大腸炎の検査方法および装置ならびに治療薬のスクリーニング方法
20日前
ダイキン工業株式会社
中空微粒子の製造方法、中空微粒子、相分離微粒子、水分散体及び組成物
2か月前
国立大学法人神戸大学
標的化したDNA配列の核酸塩基を特異的に変換するゲノム配列の改変方法及びそれに用いる分子複合体
3か月前
合同酒精株式会社
麦汁の製造方法
3か月前
池田食研株式会社
RNAの合成方法
3か月前
東洋紡株式会社
細菌からの核酸抽出法
4か月前
杏林製薬株式会社
核酸検出用PCR溶液
1か月前
個人
細胞培養容器
22日前
日本バイリーン株式会社
細胞用支持基材
27日前
サッポロビール株式会社
飲料
1か月前
東洋紡株式会社
改変型RNAポリメラーゼ
22日前
熊本県
低褐変レタスとその作製方法
4か月前
日油株式会社
蛋白質安定化剤
21日前
学校法人近畿大学
培養肉の製造方法
2か月前
東洋紡株式会社
ウイルスからの核酸抽出法
4か月前
JNC株式会社
アデノ随伴ウイルスの精製方法
2か月前
続きを見る
他の特許を見る




 特許ウォッチ
特許ウォッチ