TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025041861
公報種別
公開特許公報(A)
公開日
2025-03-26
出願番号
2024228449,2021010565
出願日
2024-12-25,2021-01-26
発明の名称
リガーゼ変異体
出願人
味の素株式会社
代理人
弁理士法人酒井国際特許事務所
主分類
C12N
9/88 20060101AFI20250318BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】優れた性質を有するリガーゼ変異体を提供すること。
【解決手段】下記(1)、(2)または(3)のリガーゼ変異体:
(1)配列番号1のアミノ酸配列に対して95%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体;
(2)配列番号2のアミノ酸配列に対して90%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体;または
(3)配列番号3のアミノ酸配列に対して97%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体。
【選択図】なし
特許請求の範囲
【請求項1】
下記(1)、(2)または(3)のリガーゼ変異体:
(1)配列番号1のアミノ酸配列に対して95%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体;
(2)配列番号2のアミノ酸配列に対して90%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体;または
(3)配列番号3のアミノ酸配列に対して97%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体。
続きを表示(約 570 文字)
【請求項2】
核酸が、DNAおよび/または修飾核酸を含んでいてもよい一本鎖RNAまたは二本鎖RNAである、請求項1記載のリガーゼ変異体。
【請求項3】
請求項1または2記載のリガーゼ変異体の存在下で核酸材料を連結して核酸生成物を生成することを含み、
核酸材料が、一本鎖核酸材料、二本鎖核酸材料、およびそれらの混合物からなる群より選ばれる、核酸生成物の製造方法。
【請求項4】
核酸材料がRNAである、請求項3記載の方法。
【請求項5】
核酸材料が4本以上の一本鎖RNAである、請求項3または4記載の方法。
【請求項6】
核酸生成物が12~27塩基長の相補部分を含む、請求項3~5のいずれか一項記載の方法。
【請求項7】
核酸材料がDNAおよび/または修飾核酸を含む、請求項3~6のいずれか一項記載の方法。
【請求項8】
核酸材料の濃度が1μM以上である、請求項3~7のいずれか一項記載の方法。
【請求項9】
核酸生成物がsiRNAである、請求項3~8のいずれか一項記載の方法。
【請求項10】
請求項1または2記載のリガーゼ変異体をコードするポリヌクレオチド。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、リガーゼ変異体などに関する。
続きを表示(約 4,200 文字)
【背景技術】
【0002】
T4 RNAリガーゼ2は、ATPの存在下でリボヌクレオチドの連結能を有するRNAリガーゼ(EC6.5.1.3)の一種であり、エシェリヒアへの感染能を有するT4バクテリオファージに由来する酵素である(NP_049790)。T4 RNAリガーゼ2は、ホスホジエステル結合を形成することにより、核酸の5’末端のリン酸基(ドナー)を3’末端のヒドロキシル基(アクセプター)に連結する能力を有する。T4 RNAリガーゼ2は、突出末端を有する二本鎖RNAの連結、および二本鎖RNA中のニックの連結等の反応に使用されている。T4 RNAリガーゼ2はまた、RNAのみならず、DNA、ならびにDNAおよびRNA以外の修飾核酸も基質として利用できることが知られている。また、T4 RNAリガーゼ2には、幾つかの変異体が知られている。T4 RNAリガーゼ2の先行技術については、下記を参照することができる。
【先行技術文献】
【特許文献】
【0003】
国際公開第2008/094599号
【非特許文献】
【0004】
[online],INTERNET,NCBI Protein Database,2018年8月13日,NP_049790,検索日:2020年2月27日,URL,https://www.ncbi.nlm.nih.gov/protein/NP_049790
Chauleau, M., & Shuman, S. (2013). Kinetic mechanism of nick sealing by T4 RNA ligase 2 and effects of 3′-OH base mispairs and damaged base lesions. RNA, 19(12): 1840-1847.
Nandakumar, J., Shuman, S., Lima, C. D. (2006). RNA ligase structures reveal the basis for RNA specificity and conformational changes that drive ligation forward. Cell, 6;127(1):71-84.
Ho, C. K., & Shuman, S. (2002). Bacteriophage T4 RNA ligase 2 (gp24.1) exemplifies a family of RNA ligases found in all phylogenetic domains. PNAS, 1;99(20):12709-14.
Nandakumar, J., Ho, C. K., Lima, C. D., Shuman, S. (2004). RNA substrate specificity and structure-guided mutational analysis of bacteriophage T4 RNA ligase 2. J. Biol. Chem., 23;279(30):31337-47.
Nandakumar, J. & Shuman, S. (2004). How an RNA ligase discriminates RNA versus DNA damage. Mol. Cell., 22;16(2):211-21.
Nandakumar, J. & Shuman, S. (2005). Dual mechanisms whereby a broken RNA end assists the catalysis of its repair by T4 RNA ligase 2. J. Biol. Chem., 24;280(25):23484-9.
Yin, S., Ho, C. K., Shuman, S. (2003). Structure-function analysis of T4 RNA ligase 2. J. Biol. Chem., 16;278(20):17601-8.
Yin, S., Kiong Ho, C., Miller, E. S., Shuman, S. (2004). Characterization of bacteriophage KVP40 and T4 RNA ligase 2. Virology, 5;319(1):141-51.
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明の目的は、優れた性質を有するリガーゼ変異体を提供することである。
【課題を解決するための手段】
【0006】
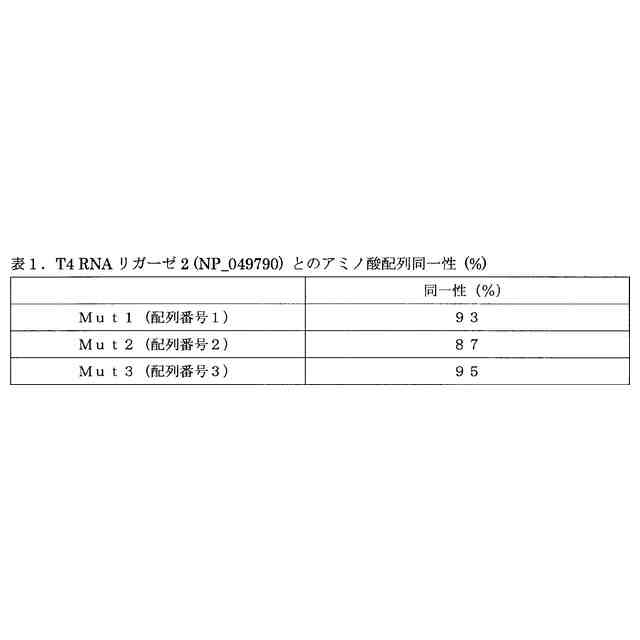
本発明者らは、鋭意検討した結果、T4 RNAリガーゼ2(NP_049790)に対してそれぞれ93%、87%および95%のアミノ酸配列同一性を示し(後述の表1を参照)、かつ当該T4 RNAリガーゼ2に比し優れた性質を有する3種のリガーゼ変異体Mut1~3(配列番号1~3)を開発することに成功し、もって本発明を完成するに至った。上記先行技術は、このような3種のリガーゼ変異体を教示も示唆もしていない。
【0007】
すなわち、本発明は、以下のとおりである。
〔1〕下記(1)、(2)または(3)のリガーゼ変異体:
(1)配列番号1のアミノ酸配列に対して95%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体;
(2)配列番号2のアミノ酸配列に対して90%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体;または
(3)配列番号3のアミノ酸配列に対して97%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体。
〔2〕核酸が、DNAおよび/または修飾核酸を含んでいてもよい一本鎖RNAまたは二本鎖RNAである、〔1〕のリガーゼ変異体。
〔3〕〔1〕または〔2〕のリガーゼ変異体の存在下で核酸材料を連結して核酸生成物を生成することを含み、
核酸材料が、一本鎖核酸材料、二本鎖核酸材料、およびそれらの混合物からなる群より選ばれる、核酸生成物の製造方法。
〔4〕核酸材料がRNAである、〔3〕の方法。
〔5〕核酸材料が4本以上の一本鎖RNAである、〔3〕または〔4〕の方法。
〔6〕核酸生成物が12~27塩基長の相補部分を含む、〔3〕~〔5〕のいずれかの方法。
〔7〕核酸材料がDNAおよび/または修飾核酸を含む、〔3〕~〔6〕のいずれかの方法。
〔8〕核酸材料の濃度が1μM以上である、〔3〕~〔7〕のいずれかの方法。
〔9〕核酸生成物がsiRNAである、〔3〕~〔8〕のいずれかの方法。
〔10〕〔1〕または〔2〕のリガーゼ変異体をコードするポリヌクレオチド。
〔11〕〔10〕のポリヌクレオチドを含む発現ベクター。
〔12〕〔1〕または〔2〕のリガーゼ変異体をコードするポリヌクレオチド、およびそれに作動可能に連結されたプロモーターを含む発現単位を含む形質転換微生物。
〔13〕〔1〕または〔2〕のリガーゼ変異体を、〔12〕の形質転換微生物を用いて生成することを含む、リガーゼ変異体の製造方法。
【発明の効果】
【0008】
本発明によれば、核酸材料から核酸生成物(例、siRNAやヘテロ二本鎖核酸等の修飾核酸)を効率的に製造することができる。
【図面の簡単な説明】
【0009】
図1は、本発明のリガーゼ変異体(Mut1~3)のアミノ酸配列(配列番号1~3)を示す図である。
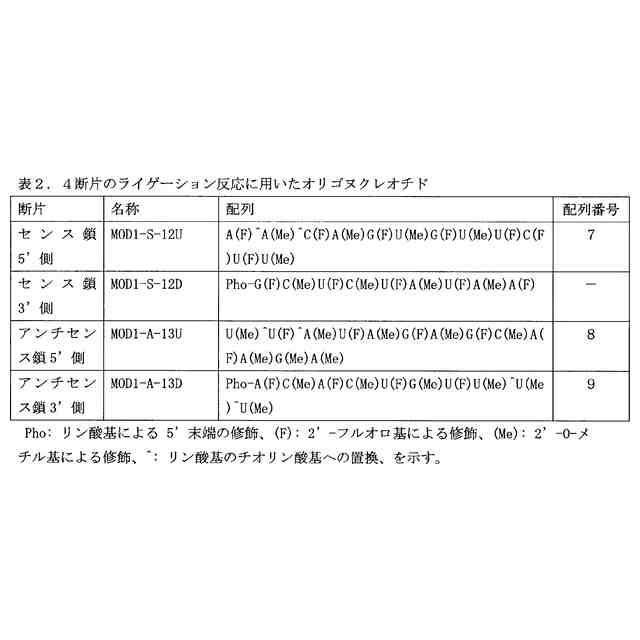
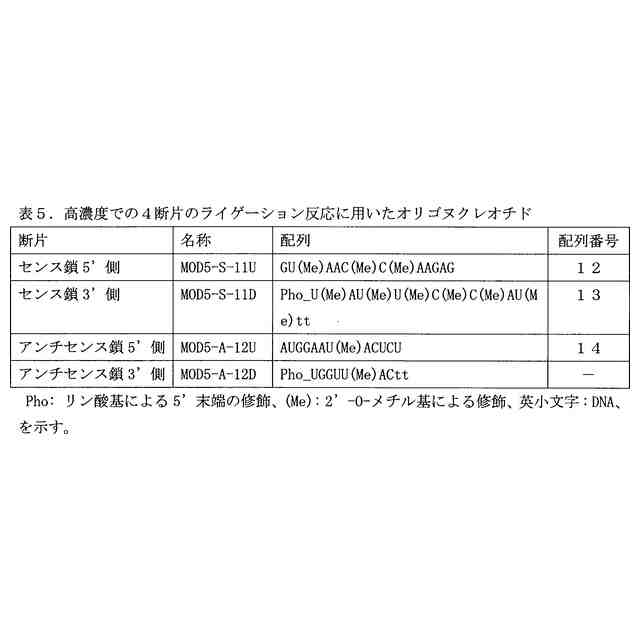
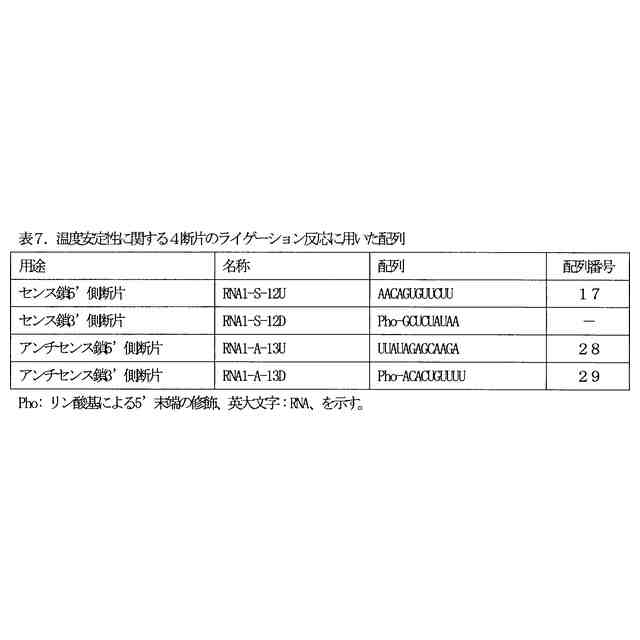
図2は、4断片の一本鎖オリゴヌクレオチドのライゲーション反応により生成する二本鎖オリゴヌクレオチドを示す図である。修飾ヌクレオチド残基の表記は、表2のものと同じである。
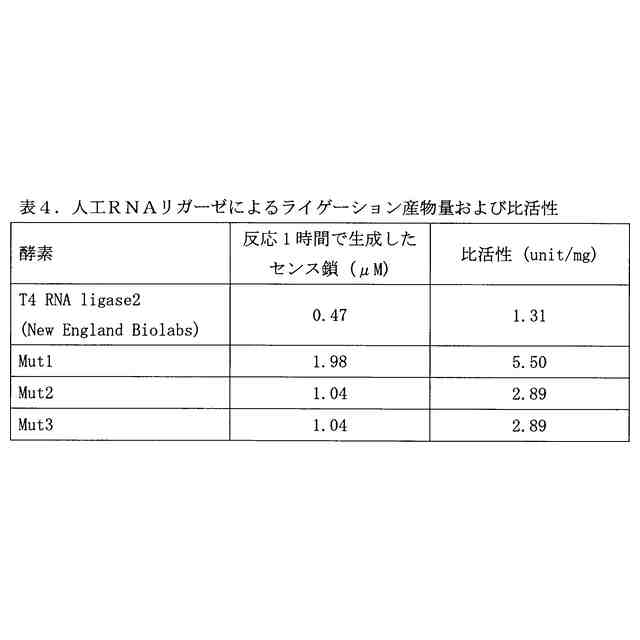
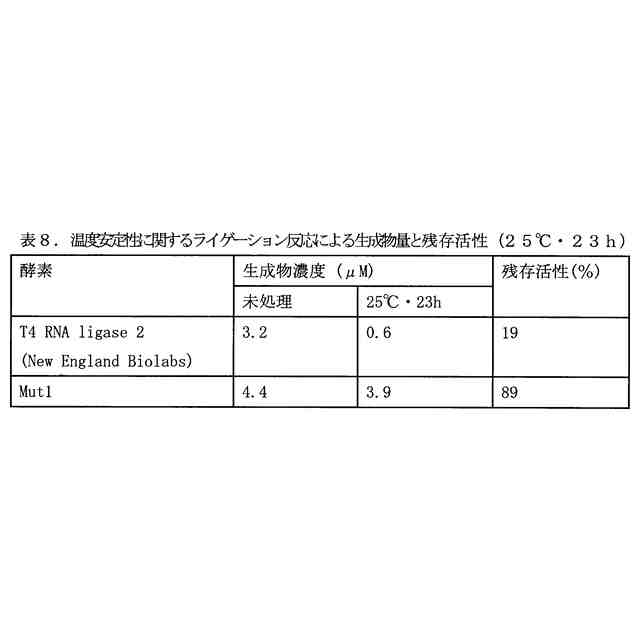
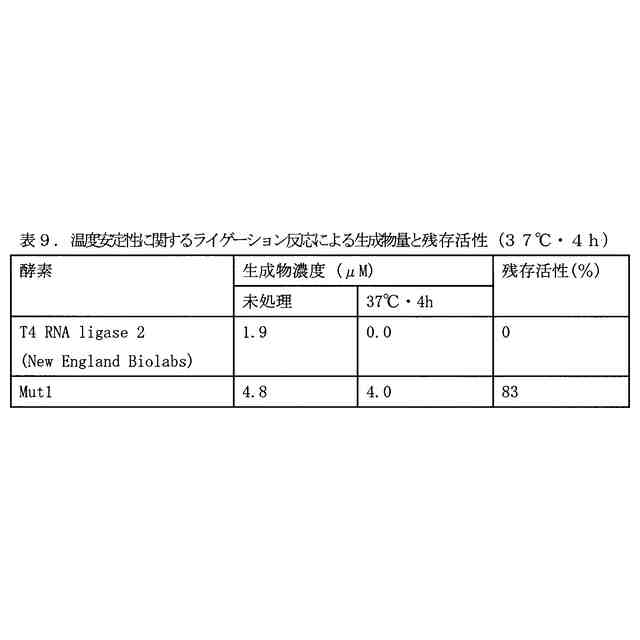
図3は、(A)4断片の一本鎖オリゴヌクレオチドのライゲーション反応により生成する二本鎖オリゴヌクレオチド、および(B)当該二本鎖オリゴヌクレオチドの生成のタイムコースを示す図である。修飾ヌクレオチド残基の表記は、表4のものと同じである。
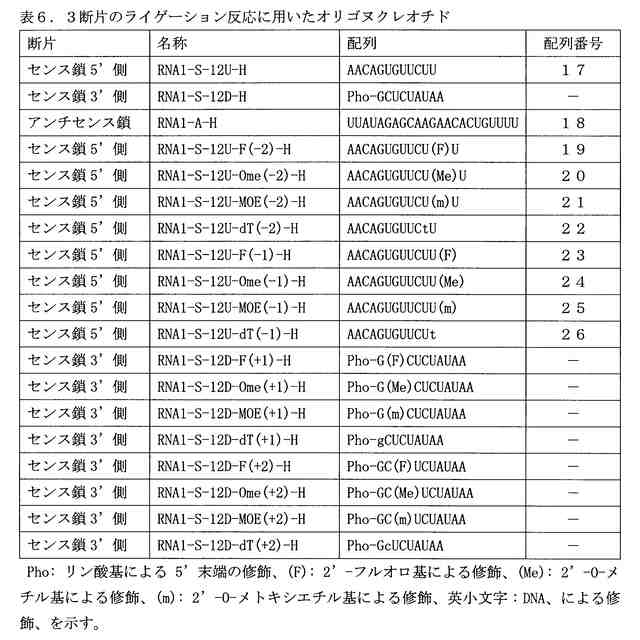
図4は、(A)3断片の一本鎖オリゴヌクレオチドのライゲーション反応により生成する二本鎖オリゴヌクレオチド、および(B)当該二本鎖オリゴヌクレオチドの量(反応時間15分)を示す図である。ライゲーションポイントから-2位、-1位、+1位、+2位のヌクレオチド残基の2’位がフッ素原子(F)、O-メチル(Ome)、O-メトキシエチル(MOE)に修飾された、または水素原子(DNA)に置換されたオリゴヌクレオチドを基質として用いた。
【発明を実施するための形態】
【0010】
1.リガーゼ変異体
本発明は、下記(1)、(2)または(3)のリガーゼ変異体を提供する:
(1)配列番号1のアミノ酸配列に対して95%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体;
(2)配列番号2のアミノ酸配列に対して90%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体;または
(3)配列番号3のアミノ酸配列に対して97%以上の同一性を示すアミノ酸配列を含み、かつ、核酸の連結活性を有するリガーゼ変異体。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
味の素株式会社
包装袋
7日前
味の素株式会社
樹脂組成物
5日前
味の素株式会社
がん関連疲労に対する抗疲労剤
4日前
味の素株式会社
電子レンジ加熱調理用調味料及びその製造方法
5日前
味の素株式会社
Nε-長鎖アシルリジン結晶の製造方法及びその結晶を含有する組成物
18日前
合同酒精株式会社
麦汁の製造方法
1か月前
JNC株式会社
アデノ随伴ウイルスの精製方法
6日前
日本特殊陶業株式会社
メタン発生抑制装置
20日前
株式会社アテクト
培養シート
19日前
住友化学株式会社
細胞チップ
1か月前
花王株式会社
肌タイプの分類方法
18日前
ニプロ株式会社
コネクタ及びバッグ
18日前
学校法人近畿大学
遺伝子組換えコクナーゼおよびその前駆体
12日前
学校法人上智学院
生物の管理方法
1か月前
国立大学法人東京科学大学
慢性腎臓病の検査方法
26日前
関西電力株式会社
微細藻類培養システム及び微細藻類培養方法
11日前
個人
分泌型タンパク質の合成方法
1か月前
横河電機株式会社
伝熱装置、加熱装置及び処理装置
4日前
国立大学法人大阪大学
発光タンパク質
19日前
エイブル株式会社
無菌コネクター
19日前
学校法人 埼玉医科大学
膜構造体、及び膜構造体の製造方法
4日前
東海漬物株式会社
新規酪酸菌とその用途
1か月前
三菱ケミカル株式会社
藻類及び油脂製造方法
1日前
住友化学株式会社
細胞接触用溶液又はそのゲル
1か月前
住友化学株式会社
細胞接触用溶液又はそのゲル
1か月前
花王株式会社
4-ヒドロキシ安息香酸類の製造方法
12日前
日本精工株式会社
実験支援システムおよび情報取得方法
19日前
公益財団法人実中研
細胞保存液及び細胞保存方法
5日前
浜松ホトニクス株式会社
ルシフェラーゼ-ペプチド連結体
22日前
公立大学法人横浜市立大学
リピート伸長病の検出方法
7日前
国立大学法人東海国立大学機構
卵巣癌を検査する方法
11日前
株式会社神鋼環境ソリューション
メタン発酵槽、及び、メタン発酵槽の運転方法
7日前
株式会社日立製作所
塩基検出用長鎖ssDNAプライマー
1か月前
株式会社日立製作所
細胞剥離装置および細胞剥離方法
19日前
個人
卵巣癌の予後を判定するためのバイオマーカー、判定方法、および治療薬
6日前
国立大学法人東北大学
ペプチド又はその塩、ならびに核酸
1か月前
続きを見る
他の特許を見る

 特許ウォッチ
特許ウォッチ