TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
公開番号
2025069623
公報種別
公開特許公報(A)
公開日
2025-05-01
出願番号
2023179446
出願日
2023-10-18
発明の名称
発光タンパク質
出願人
国立大学法人大阪大学
代理人
弁理士法人平木国際特許事務所
主分類
C12N
15/62 20060101AFI20250423BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】複数の波長域で高光度に発光する、発光タンパク質を提供する。
【解決手段】バクテリアルシフェラーゼと蛍光タンパク質との複合体を含み、前記バクテリアルシフェラーゼはLuxAサブユニット及びLuxBサブユニットを有し、前記蛍光タンパク質はLuxAサブユニットを介してバクテリアルシフェラーゼと融合している、発光タンパク質。
【選択図】なし
特許請求の範囲
【請求項1】
バクテリアルシフェラーゼと蛍光タンパク質との複合体を含み、前記バクテリアルシフェラーゼはLuxAサブユニット及びLuxBサブユニットを有し、前記蛍光タンパク質はLuxAサブユニットと融合している、発光タンパク質。
続きを表示(約 1,400 文字)
【請求項2】
前記LuxAサブユニットが、配列番号1で表されるアミノ酸配列からなるポリペプチド又は配列番号1で表されるアミノ酸配列と50%以上の配列同一性を有するアミノ酸配列からなるポリペプチドである、請求項1に記載の発光タンパク質。
【請求項3】
前記LuxBサブユニットが、配列番号2で表されるアミノ酸配列からなるポリペプチド又は配列番号2で表されるアミノ酸配列と50%以上の配列同一性を有するアミノ酸配列からなるポリペプチドである、請求項1に記載の発光タンパク質。
【請求項4】
前記蛍光タンパク質が、シアン蛍光タンパク質、緑色蛍光タンパク質、黄色蛍光タンパク質、橙色蛍光タンパク質及び赤色蛍光タンパク質から選択される、請求項1に記載の発光タンパク質。
【請求項5】
前記シアン蛍光タンパク質が、配列番号3で表されるアミノ酸配列からなるポリペプチド、若しくは配列番号3で表されるアミノ酸配列と60%以上の配列同一性を有し、かつ、極大蛍光波長が450~500nmにある蛍光を発する機能を有するポリペプチド、又はこれらのいずれかのポリペプチドの循環置換体である;
前記緑色蛍光タンパク質が、配列番号4で表されるアミノ酸配列からなるポリペプチド、若しくは配列番号4で表されるアミノ酸配列と60%以上の配列同一性を有し、かつ、極大蛍光波長が500~520nmにある蛍光を発する機能を有するポリペプチド、又はこれらのいずれかのポリペプチドの循環置換体である;
前記黄色蛍光タンパク質が、配列番号5で表されるアミノ酸配列からなるポリペプチド、若しくは配列番号5で表されるアミノ酸配列と60%以上の配列同一性を有し、かつ、極大蛍光波長が520~550nmにある蛍光を発する機能を有するポリペプチド、又はこれらのいずれかのポリペプチドの循環置換体である;
前記橙色蛍光タンパク質が、配列番号6で表されるアミノ酸配列からなるポリペプチド、若しくは配列番号6で表されるアミノ酸配列と60%以上の配列同一性を有し、かつ、極大蛍光波長が550~580nmにある蛍光を発する機能を有するポリペプチド、又はこれらのいずれかのポリペプチドの循環置換体である;又は、
前記赤色蛍光タンパク質が、配列番号7で表されるアミノ酸配列からなるポリペプチド、若しくは配列番号7で表されるアミノ酸配列と60%以上の配列同一性を有し、かつ、極大蛍光波長が580~620nmにある蛍光を発する機能を有するポリペプチド、又はこれらのいずれかのポリペプチドの循環置換体である、
請求項4に記載の発光タンパク質。
【請求項6】
バクテリアルシフェラーゼのLuxAと蛍光タンパク質との融合体をコードする塩基配列を含む、核酸。
【請求項7】
さらに、バクテリアルシフェラーゼのLuxBをコードする塩基配列を含む、請求項6に記載の核酸。
【請求項8】
請求項6に記載の核酸が組み込まれた、発現用ベクター。
【請求項9】
バクテリアルシフェラーゼのLuxAと蛍光タンパク質との融合体をコードする塩基配列を有する核酸、及びLuxBをコードする塩基配列を有する核酸が導入された、細胞。
【請求項10】
細菌、真菌細胞、植物細胞又は動物細胞である、請求項9に記載の細胞。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、発光バクテリア由来のルシフェラーゼを含む発光タンパク質に関する。
続きを表示(約 4,200 文字)
【背景技術】
【0002】
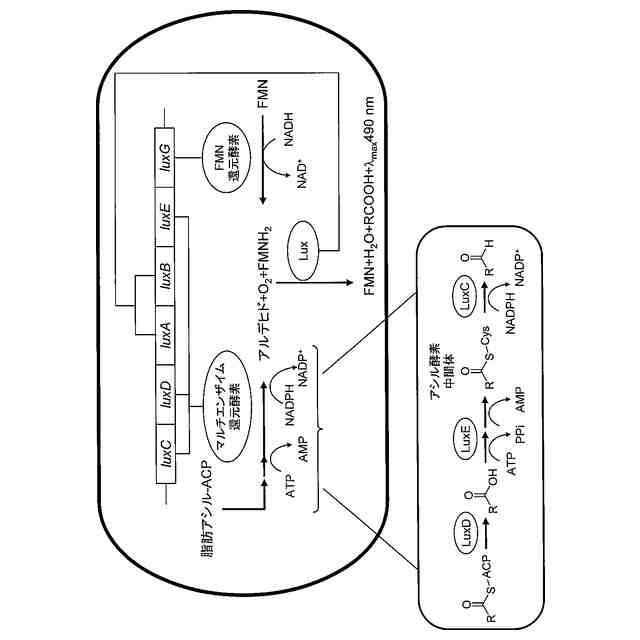
ルシフェラーゼと呼ばれる触媒酵素により発光基質が酸化される発光現象のうち、生物内にて起こるものは生物発光と分類される。生物発光を示す生物は数千種類知られており、その中には、ルシフェラーゼ及び発光基質が同定されており、これらは、例えば、遺伝子工学にて光を指標として細胞中のタンパク質等を定量する目的で使用されている。
【0003】
真正細菌(細菌、バクテリア)には、発光を生じる種(発光バクテリア)が複数存在する。発光バクテリアにおける光生成は、バクテリア由来のルシフェラーゼによって触媒される、基質の酵素反応によって生じる(非特許文献1及び2)。バクテリア由来のルシフェラーゼは、具体的には、還元フラビンモノヌクレオチド(FMNH
2
)及び長鎖脂肪アルデヒド(RCHO)を酸化して、フラビンモノヌクレオチド(FMN)及び対応する長鎖脂肪酸(RCOOH)を生成するが、この反応時に、490nm付近に極大波長を有する光が発生する。
【0004】
発光バクテリアの発光に必要な基本的な酵素は、luxCDABEという単一のオペロン(luxオペロン)によってコードされており、このオペロンは、発光バクテリアのすべての種に見られる(非特許文献1及び2)。luxA及びluxBは、それぞれ、バクテリアルシフェラーゼのヘテロ二量体タンパク質のα(LuxA)サブユニット及びβ(LuxB)サブユニットをコードする。luxC、luxD及びluxEは、ルシフェラーゼの基質である脂肪アルデヒドを合成し、再循環させるための複合体をコードする遺伝子である。発光バクテリア以外の細菌又は酵母細胞で、上記の5つのlux遺伝子を共発現することにより、外部からの基質の供給を要することなく、自発的な発光現象(自発光)を示すことが観察された(非特許文献3及び4)。哺乳動物細胞又は植物細胞においては、上記の5つのlux遺伝子に加えて、FMNオキシドレダクターゼ(LuxG)を共発現させることで、十分な量のFMNH
2
を生成することができ、これにより、安定な生物発光が可能となる(非特許文献5-8)。
【0005】
生物発光イメージングは、蛍光タンパク質によるイメージングとは異なり、外部からの励起光を必要としないため、光毒性、光退色、試料由来の蛍光バックグラウンド等の問題が活性し難い、という利点を有する。特に、バクテリアルシフェラーゼに基づくレポーターは、シグナル対ノイズ比(S/N比)が高く、かつ取り扱いが容易である、という利点も有する。また、励起光のスペクトル分析への干渉が生じないため、蛍光タンパク質よりも広いスペクトル範囲の解析が可能である。一方で、バクテリア由来の生物発光イメージングは、低輝度である、露光時間が長い、といった課題があり、その利用が制限されてきた(非特許文献9及び10)。
【0006】
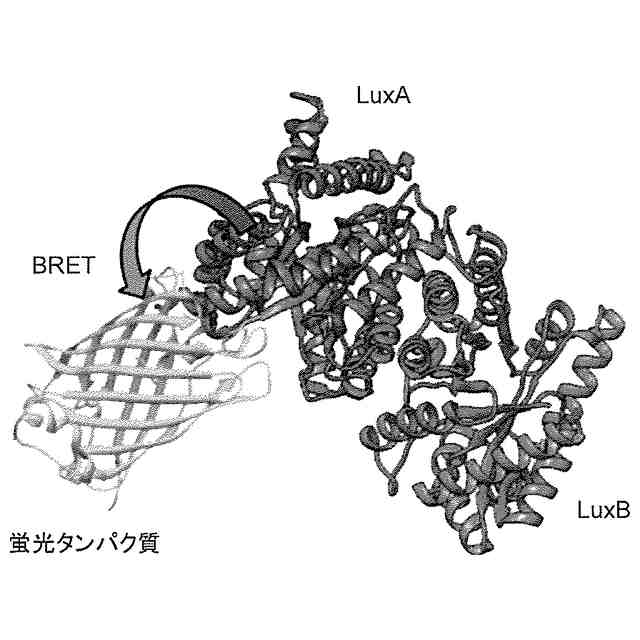
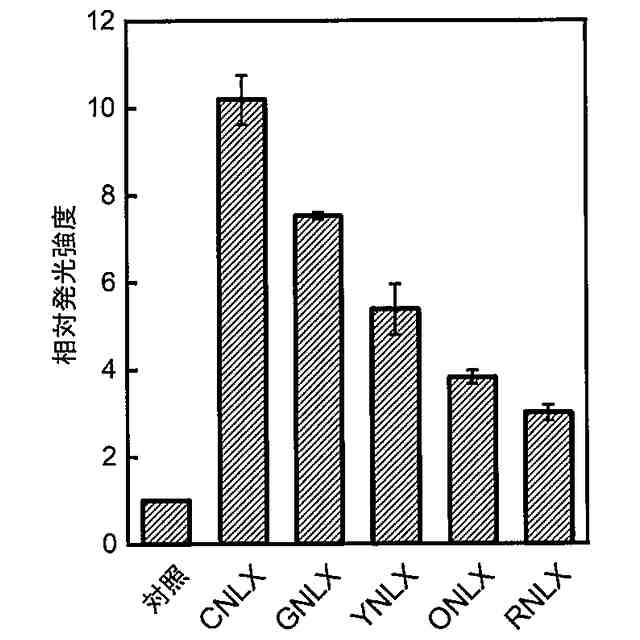
非特許文献11には、バクテリア由来のLuxBと円順列変異を導入した黄色蛍光タンパク質Venusとを融合することにより、細胞内で生物発光共鳴エネルギー移動(bioluminescence resonance energy transfer(BRET))を生じさせ、Lux単独と比較して、短時間で10倍もの発光強度を達成できたことが記載されている。さらに、これを植物細胞及び動物細胞に導入することで、これらの細胞においても高光度化を達成したことも明らかにされた。
【先行技術文献】
【非特許文献】
【0007】
Meighen, E., A., Microbiol. Rev. 55, 123-142 (1991)
Dunlap, P., Adv. Biochem. Eng. Biotechnol. 144, 37-64 (2014)
Frackman, S. et al., J. Bacteriol. 172, 5767-5773 (1990)
Gupta, R. K. et al., FEMS Yeast Res. 4, 305-313 (2003)
Close, D. M. et al., PLoS One 5, e12441 (2010)
Krichevsky, A. et al., PLoS One 5, e15461 (2010)
Xu, T. et al., PLoS One 9, e96347 (2014)
Gregor, C. et al. Proc. Natl. Acad. Sci. USA 116, 26491-26496 (2019)
Close, D. M. et al., J. Biomed. Opt. 16, 047003 (2011)
Gregor, C. et al., Proc. Natl. Acad. Sci. USA 115, 962-967 (2018)
Kaku, T. et al., Nature Scientific Reports, 11, 14994 (2021)
【発明の概要】
【発明が解決しようとする課題】
【0008】
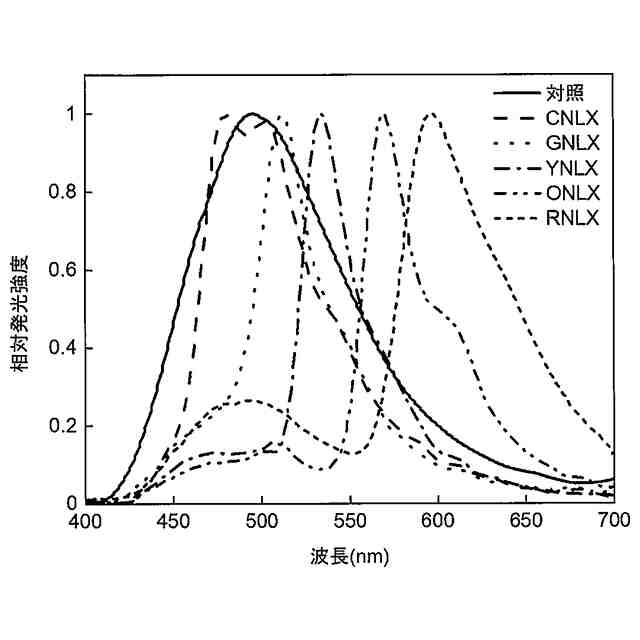
上記の通り、バクテリア由来のluxオペロンを利用し、さらに蛍光タンパク質VenusをluxBと融合させることで、細胞内において高光度で自発光可能な発光タンパク質を構築できることが示された。この系で発生する光は、黄緑色(極大波長528nm)であるが、複数種のレポーターを要する細胞実験等に使用するには、異なる波長の光を発する複数種の発光タンパク質の存在が望まれる。
【0009】
本発明は、複数の波長域で高光度に発光する、発光タンパク質を提供することを目的とする。また、本発明は、複数の波長域で高光度に発光する発光タンパク質について、これをコードする核酸、該核酸を組み込んだベクター、及び該核酸を組み込んだ細胞を提供することを目的とする。
【課題を解決するための手段】
【0010】
本発明は、以下を提供する。
[1]バクテリアルシフェラーゼと蛍光タンパク質との複合体を含み、前記バクテリアルシフェラーゼはLuxAサブユニット及びLuxBサブユニットを有し、前記蛍光タンパク質はLuxAサブユニットと融合している、発光タンパク質。
[2]前記LuxAサブユニットが、配列番号1で表されるアミノ酸配列からなるポリペプチド、又は配列番号1で表されるアミノ酸配列と50%以上の配列同一性を有するアミノ酸配列からなるポリペプチドである、[1]に記載の発光タンパク質。
[3]前記LuxBサブユニットが、配列番号2で表されるアミノ酸配列からなるポリペプチド又は配列番号2で表されるアミノ酸配列と50%以上の配列同一性を有するアミノ酸配列からなるポリペプチドである、[1]又は[2]に記載の発光タンパク質。
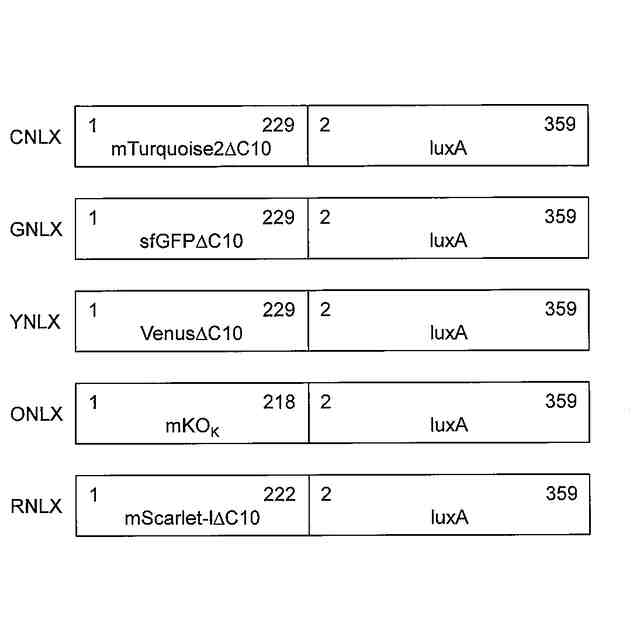
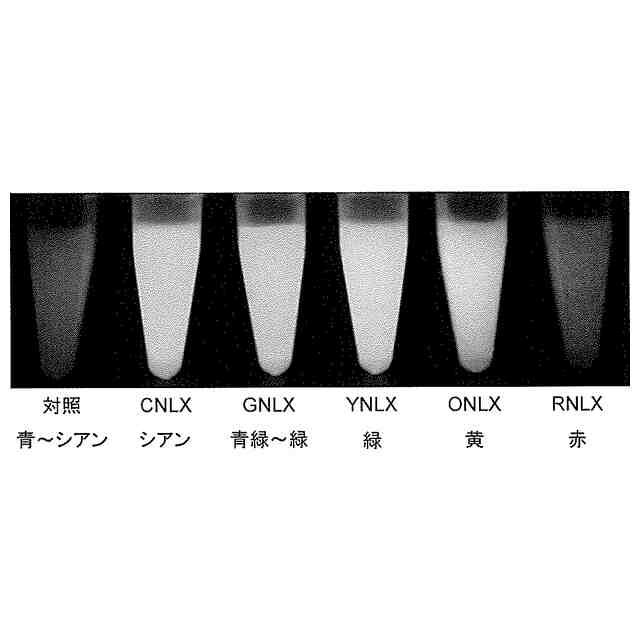
[4]前記蛍光タンパク質が、シアン蛍光タンパク質、緑色蛍光タンパク質、黄色蛍光タンパク質、橙色蛍光タンパク質及び赤色蛍光タンパク質から選択される、[1]~[3]のいずれかに記載の発光タンパク質。
[5]前記シアン蛍光タンパク質が、配列番号3で表されるアミノ酸配列からなるポリペプチド、若しくは配列番号3で表されるアミノ酸配列と60%以上の配列同一性を有し、かつ、極大蛍光波長が450~500nmにある蛍光を発する機能を有するポリペプチド、又はこれらのいずれかのポリペプチドの循環置換体である;前記緑色蛍光タンパク質が、配列番号4で表されるアミノ酸配列からなるポリペプチド、若しくは配列番号4で表されるアミノ酸配列と60%以上の配列同一性を有し、かつ、極大蛍光波長が500~520nmにある蛍光を発する機能を有するポリペプチド、又はこれらのいずれかのポリペプチドの循環置換体である;前記黄色蛍光タンパク質が、配列番号5で表されるアミノ酸配列からなるポリペプチド、若しくは配列番号5で表されるアミノ酸配列と60%以上の配列同一性を有し、かつ、極大蛍光波長が520~550nmにある蛍光を発する機能を有するポリペプチド、又はこれらのいずれかのポリペプチドの循環置換体である;前記橙色蛍光タンパク質が、配列番号6で表されるアミノ酸配列からなるポリペプチド、若しくは配列番号6で表されるアミノ酸配列と60%以上の配列同一性を有し、かつ、極大蛍光波長が550~580nmにある蛍光を発する機能を有するポリペプチド、又はこれらのいずれかのポリペプチドの循環置換体である;又は、前記赤色蛍光タンパク質が、配列番号7で表されるアミノ酸配列からなるポリペプチド、若しくは配列番号7で表されるアミノ酸配列と60%以上の配列同一性を有し、かつ、極大蛍光波長が580~620nmにある蛍光を発する機能を有するポリペプチド、又はこれらのいずれかのポリペプチドの循環置換体である、[4]に記載の発光タンパク質。
[6]バクテリアルシフェラーゼのLuxAと蛍光タンパク質との融合体をコードする塩基配列を含む、核酸。
[7]さらに、バクテリアルシフェラーゼのLuxBをコードする塩基配列を含む、[6]に記載の核酸。
[8][6]又は[7]に記載の核酸が組み込まれた、発現用ベクター。
[9]バクテリアルシフェラーゼのLuxAと蛍光タンパク質との融合体をコードする塩基配列を有する核酸、及びLuxBをコードする塩基配列を有する核酸が導入された、細胞。
[10]細菌、真菌細胞、植物細胞又は動物細胞である、[9]に記載の細胞。
[11]バクテリアルシフェラーゼのLuxAと蛍光タンパク質との融合体をコードする塩基配列を有する核酸、及びLuxBをコードする塩基配列を有する核酸を発現させることを含む、発光タンパク質の製造方法。
【発明の効果】
(【0011】以降は省略されています)
この特許をJ-PlatPat(特許庁公式サイト)で参照する
関連特許
国立大学法人大阪大学
滅菌器
今日
国立大学法人大阪大学
モータ装置
2か月前
国立大学法人大阪大学
細胞シート
5か月前
国立大学法人大阪大学
歩行訓練装置
2日前
国立大学法人大阪大学
発光タンパク質
7か月前
国立大学法人大阪大学
半月板の治療薬
2か月前
国立大学法人大阪大学
メタン反応装置
今日
国立大学法人大阪大学
原子力発電装置
6か月前
国立大学法人大阪大学
眼瞼痙攣緩和具
4か月前
国立大学法人大阪大学
内視鏡システム
6か月前
国立大学法人大阪大学
ロボットハンド
3か月前
国立大学法人大阪大学
漏洩情報抑制回路
4か月前
国立大学法人大阪大学
動体視力向上用組成物
29日前
国立大学法人大阪大学
試料中の標的物質の検出
2か月前
国立大学法人大阪大学
化学反応方法及び反応装置
2か月前
国立大学法人大阪大学
溶接装置および積層造形装置
今日
トヨタ自動車株式会社
電池
5か月前
国立大学法人大阪大学
電気泳動装置および電気泳動法
3か月前
国立大学法人大阪大学
音声分析方法および音声分析装置
今日
国立大学法人大阪大学
コネクタ取付具およびコネクタ部材
7か月前
大和ハウス工業株式会社
反応装置
2か月前
大和ハウス工業株式会社
反応装置
2か月前
大和ハウス工業株式会社
反応装置
6か月前
国立大学法人大阪大学
CTAT電流生成回路及びBGR回路
1か月前
国立大学法人大阪大学
化合物の製造方法、及び金属多核錯体
3か月前
川崎重工業株式会社
ゼオライト触媒
2か月前
日本電信電話株式会社
量子鍵配送装置
7か月前
国立大学法人大阪大学
光発電システムの出力電力特性の最適化
7か月前
国立大学法人大阪大学
タンパク質間相互作用を可視化する方法
3か月前
国立大学法人大阪大学
溶融試料浮遊装置および溶融試料浮遊方法
2か月前
株式会社神戸製鋼所
ねじり疲労評価方法
6か月前
国立大学法人大阪大学
ナノダイヤモンドの製造方法および製造装置
5か月前
株式会社トクヤマ
ケトン誘導体の製造方法
4か月前
株式会社トクヤマ
ケトン誘導体の製造方法
1か月前
セイコーグループ株式会社
生体電位計測装置
2か月前
セイコーグループ株式会社
生体電位計測装置
2か月前
続きを見る
他の特許を見る


 特許ウォッチ
特許ウォッチ