TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025012122
公報種別
公開特許公報(A)
公開日
2025-01-24
出願番号
2023114712
出願日
2023-07-12
発明の名称
遺伝子組換えチョウ目昆虫
出願人
国立研究開発法人農業・食品産業技術総合研究機構
代理人
弁理士法人平木国際特許事務所
主分類
A01K
67/60 20250101AFI20250117BHJP(農業;林業;畜産;狩猟;捕獲;漁業)
要約
【課題】カイコ等のチョウ目昆虫において目的タンパク質を安定的かつ大量に生産するための新たな発現系を提供することを課題とする。また、目的タンパク質と融合したシルクタンパク質を含む機能性シルクを生産するための新たな方法を提供することを課題とする。
【解決手段】遺伝子組換えチョウ目昆虫であって、内在性遺伝子のシグナルペプチド又はその機能性断片をコードするエクソン配列において、目的のタンパク質又はその断片をコードする目的遺伝子配列を含み、前記目的のタンパク質又はその断片は、前記シグナルペプチド又はその機能性断片のC末端側に融合される、前記遺伝子組換えチョウ目昆虫を提供することを提供する。また、遺伝子組換えチョウ目昆虫であって、内在性遺伝子のシグナルペプチド又はその機能性断片をコードするエクソン配列において、目的のタンパク質又はその断片をコードする目的遺伝子配列を含み、前記目的のタンパク質又はその断片は、前記シグナルペプチド又はその機能性断片と、前記内在性遺伝子によってコードされる前駆体タンパク質から前記シグナルペプチドが切断されてなる成熟型タンパク質又はそのC末端断片との間に融合される、前記遺伝子組換えチョウ目昆虫を提供する。
【選択図】なし
特許請求の範囲
【請求項1】
遺伝子組換えチョウ目昆虫であって、
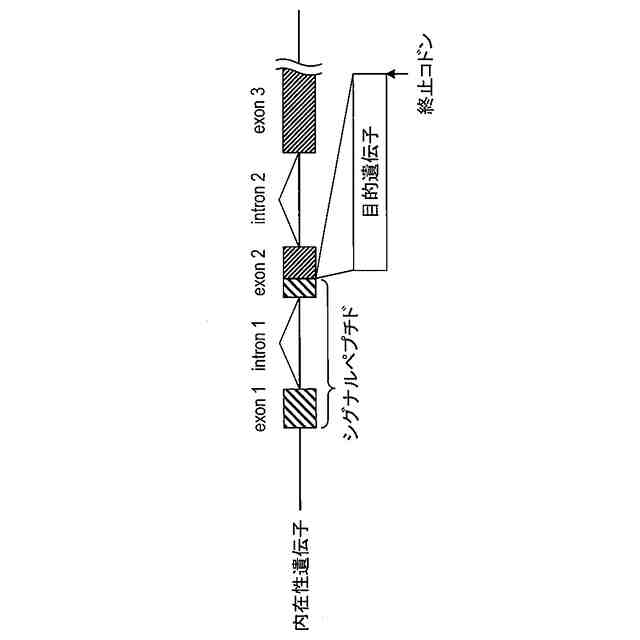
内在性遺伝子のシグナルペプチド又はその機能性断片をコードするエクソン配列において、目的のタンパク質又はその断片をコードする目的遺伝子配列を含み、
前記目的のタンパク質又はその断片は、前記シグナルペプチド又はその機能性断片と、前記内在性遺伝子によってコードされる前駆体タンパク質から前記シグナルペプチドが切断されてなる成熟型タンパク質又はそのC末端断片との間に融合される、前記遺伝子組換えチョウ目昆虫。
続きを表示(約 1,800 文字)
【請求項2】
前記成熟型タンパク質が、フィブロイン、セリシン、及び/又はフィブロヘキサマリンである、請求項1に記載の遺伝子組換えチョウ目昆虫。
【請求項3】
前記フィブロインが、フィブロインH鎖及び/又はフィブロインL鎖である、請求項2に記載の遺伝子組換えチョウ目昆虫。
【請求項4】
前記目的のタンパク質が、蛍光タンパク質、抗体、抗原ポリペプチド、酵素、サイトカイン、及び抗菌ポリペプチドからなる群から選択される、請求項1に記載の遺伝子組換えチョウ目昆虫。
【請求項5】
前記目的遺伝子配列を含む前記エクソン配列をホモ接合で含む、請求項1に記載の遺伝子組換えチョウ目昆虫。
【請求項6】
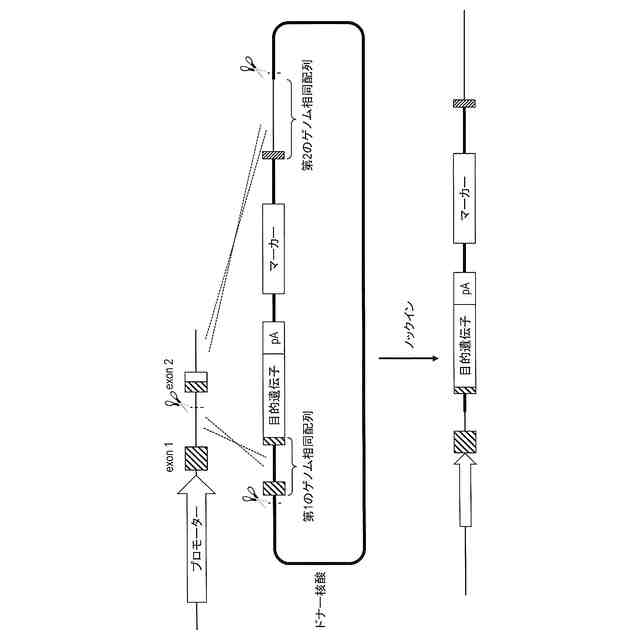
遺伝子組換えチョウ目昆虫を相同組換え法を用いて作出するためのドナー核酸であって、
前記相同組換え法は、内在性遺伝子においてイントロン配列内のゲノム切断位置をゲノム編集酵素で切断することを含み、
前記ドナー核酸は、
(a)前記内在性遺伝子に由来する、第1のゲノム相同配列及び第2のゲノム相同配列、並びに
(b)その間に配置された目的遺伝子配列
を含み、
前記第1のゲノム相同配列は、ゲノム上で前記ゲノム切断位置より5'末端側に位置する塩基から、前記イントロン配列の3'末端側に位置するエクソン配列又はその部分配列までのゲノム配列に対して相同な塩基配列からなり、かつ前記ゲノム編集酵素の認識配列に変異を有し、
前記第2のゲノム相同配列は、ゲノム上で前記エクソン配列又はその部分配列の3'末端側に位置するゲノム配列に対して相同な塩基配列からなり、
前記目的遺伝子配列は、前記内在性遺伝子のシグナルペプチド又はその機能性断片と、前記内在性遺伝子によってコードされる前駆体タンパク質から前記シグナルペプチドが切断されてなる成熟型タンパク質又はそのC末端断片との間に融合される目的のタンパク質又はその断片をコードする、前記ドナー核酸。
【請求項7】
遺伝子組換えチョウ目昆虫を相同組換え法を用いて作出するためのドナー核酸であって、
前記相同組換え法は、内在性遺伝子においてイントロン配列内のゲノム切断位置をゲノム編集酵素で切断することを含み、
前記ドナー核酸は、
(a)前記内在性遺伝子に由来する、第1のゲノム相同配列及び第2のゲノム相同配列、並びに
(b)その間に配置された目的遺伝子配列
を含み、
前記第1のゲノム相同配列は、前記イントロン配列より5'末端側に位置する塩基から前記イントロン配列の5'末端側に位置するエクソン配列又はその部分配列までのゲノム配列に対して相同な塩基配列からなり、
前記第2のゲノム相同配列は、前記エクソン配列又はその部分配列より3'末端側かつ前記ゲノム切断位置より5'末端側に位置する塩基から、前記ゲノム切断位置より3'末端側に位置する塩基までのゲノム配列に対して相同な塩基配列からなり、かつ前記ゲノム編集酵素の認識配列に変異を有し、
前記目的遺伝子配列は、前記内在性遺伝子のシグナルペプチド又はその機能性断片と、前記内在性遺伝子によってコードされる前駆体タンパク質から前記シグナルペプチドが切断されてなる成熟型タンパク質又はそのC末端断片との間に融合される目的のタンパク質又はその断片をコードする、前記ドナー核酸。
【請求項8】
前記第1のゲノム相同配列及び/又は前記第2のゲノム相同配列の前記目的遺伝子配列とは反対側の末端に、ヌクレアーゼ認識配列を含む、請求項6又は7に記載のドナー核酸。
【請求項9】
前記ヌクレアーゼ認識配列が、前記ゲノム編集酵素の認識配列、又は制限酵素認識配列である、請求項8に記載のドナー核酸。
【請求項10】
遺伝子組換えチョウ目昆虫の作出方法であって、
請求項6又は7に記載のドナー核酸、及び
前記ゲノム編集酵素又は前記ゲノム編集酵素を発現可能な状態でコードする核酸
をチョウ目昆虫の卵にマイクロインジェクション法で導入する導入工程
を含む、前記方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、遺伝子組換えチョウ目昆虫及びその作出方法等に関する。
続きを表示(約 2,100 文字)
【背景技術】
【0002】
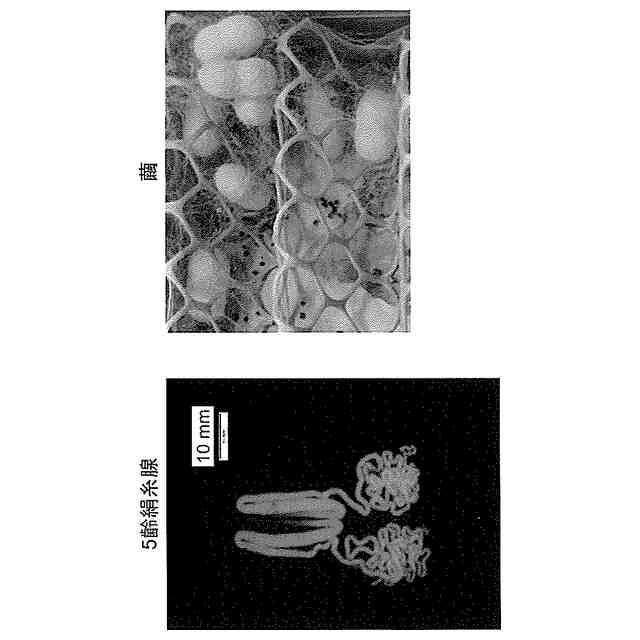
カイコ(Bombyx mori)の絹糸腺は、大量のタンパク質を短期間に合成できる能力を有している。また、カイコの絹糸腺は大型器官であるため摘出が容易であり、合成されたタンパク質は絹糸腺内腔に貯蔵されることから回収しやすいという利点もある。そのため、絹糸腺で目的のタンパク質を発現する遺伝子組換えカイコは、タンパク質の大量生産系として有望視されている。
【0003】
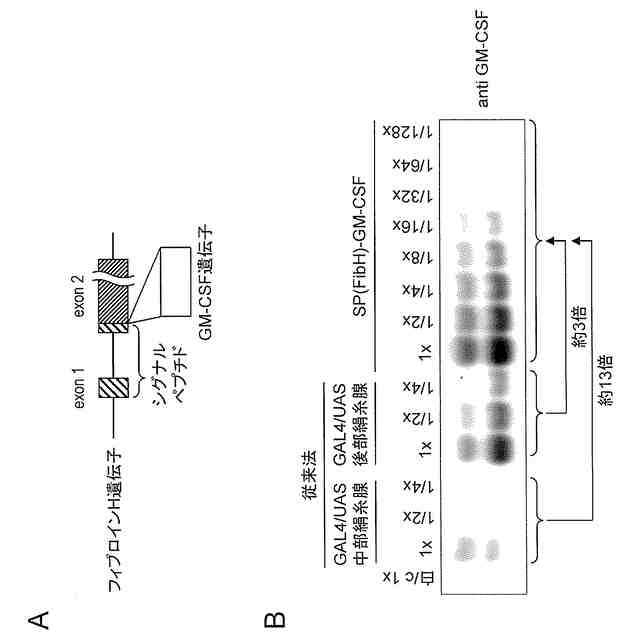
カイコの絹糸腺は、左右1対の器官であり、それぞれは、前部絹糸腺、中部絹糸腺、及び後部絹糸腺の3つの領域で構成されている。後部絹糸腺細胞内では、絹糸の繊維成分であるフィブロインを構成する3つの主要なタンパク質、フィブロインH鎖(以下、しばしば「Fib H」と略称する)、フィブロインL鎖(以下、しばしば「Fib L」と略称する)、及びフィブロヘキサマリン(p25/FHXとも呼ばれる)が発現している。また中部絹糸腺細胞内では絹糸の被覆成分であるゼラチン様タンパク質のセリシンが発現している。後部絹糸腺細胞内で発現した前記3つのタンパク質は、Fib H:Fib L:p25=6:6:1の比率で複合体(silk fibroin elementary unit;SFEU複合体)を形成し、後部絹糸腺内腔中に分泌される。これに対して、セリシンは、発現後に中部絹糸腺内腔中に分泌される。後部絹糸腺内腔中に分泌されたフィブロインは、その後、中部絹糸腺内腔に移行し、セリシンで被覆されて絹糸として吐糸される(非特許文献1)。したがって、カイコ絹糸腺をタンパク質発現系として利用する場合、中部絹糸腺又は後部絹糸腺で特異的に発現する遺伝子発現系を利用すればよい。
【0004】
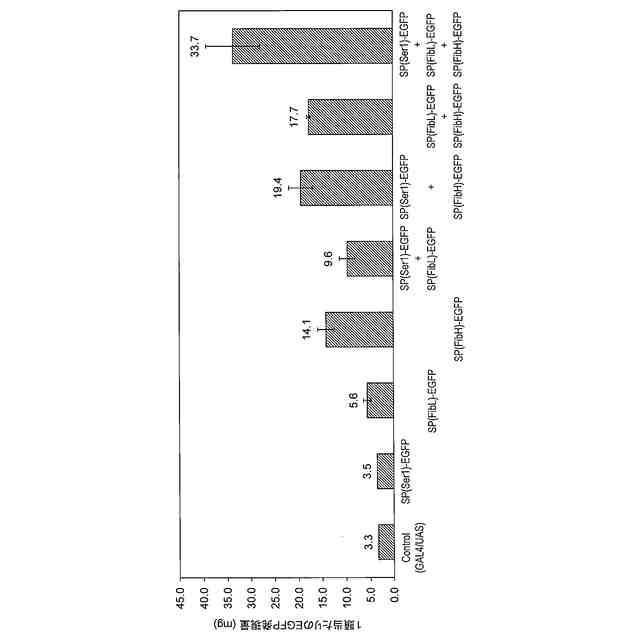
カイコ絹糸腺をタンパク質発現系として利用する場合、これまでに組換えタンパク質発現システムとしてGAL4/UASシステム(非特許文献2)、及びセリシン1プロモーター及びHr3エンハンサーを組み合わせた系による大量発現方法(非特許文献3)が報告されているが、現状ではタンパク質発現量の点で優れたGAL4/UASシステムが多く利用されている。
【0005】
GAL4/UASシステムは、酵母に由来する転写因子GAL4と制御配列UASとを組み合わせて利用する遺伝子制御システムである。カイコの絹糸腺においてタンパク質生産系として用いるGAL4/UASシステムでは、中部絹糸腺又は後部絹糸腺で特異的に発現する遺伝子のプロモーター制御下でGAL4遺伝子を発現するGAL4系統と、制御配列UASの制御下で目的タンパク質遺伝子を発現するUAS系統とをpiggyBacを用いた遺伝子組換えにより独立に樹立した後、両系統を交配することにより、絹糸腺で目的タンパク質を発現する発現系が構築される。
【0006】
GAL4/UASシステムでは、GAL4系統とUAS系統を別々に樹立した後で交配する必要があるため発現系の構築に時間を要する点や、GAL4遺伝子及びUAS制御配列はゲノム上のランダムな位置に導入されるため目的タンパク質の発現量が変動し得る点が、新たなGAL4系統及びUAS系統を樹立する際の障害となっている。また、GAL4/UASシステムでの発現量は、技術的に限界に達していると考えられる。
【0007】
非特許文献4には、カイコの絹糸腺において遺伝子組換えタンパク質を発現させるために過去に実施された様々な方法が開示されている。しかしながら、過去に報告された方法では、融合タンパク質の発現量が低いことやシルクタンパク質と融合したタンパク質の活性が失われるといった問題があった。
【0008】
したがって、目的タンパク質を安定的かつ大量に生産するための新たな方法や、目的タンパク質と融合したシルクタンパク質を効率よく生産するための新たな方法が求められている。
【先行技術文献】
【非特許文献】
【0009】
Inoue S. et al., 2000, The Journal of Biological Chemistry, 275 (51): 40517-40528.
Tatematsu K. et al., 2010, Transgenic Research, 19(3):473-87.
Tomita M. et al., 2007, Transgenic Research, 16 (4):449-465.
Tomita M. et al., 2011, Biotechnol Lett, 33:645-654.
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明の目的は、カイコ等のチョウ目昆虫において目的タンパク質を安定的かつ大量に生産するための新たな発現系を提供することである。また、目的タンパク質と融合したシルクタンパク質を含む機能性シルクを生産するための新たな方法を提供することを課題とする。
【課題を解決するための手段】
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
個人
ペット用玩具
7日前
タキイ種苗株式会社
レタス植物
今日
個人
根掛りしないイカ用疑似餌
10日前
みのる産業株式会社
移植装置
7日前
個人
ルアー
今日
株式会社東海化成
ポット保持具
6日前
AGRIST株式会社
収穫装置
3日前
株式会社クボタケミックス
弁装置
7日前
株式会社ササキコーポレーション
農作業機
7日前
大日本除蟲菊株式会社
薬剤揮散装置
今日
南京大学
藻類バイオフィルムの高速成膜方法
10日前
株式会社デンソー
潅水システム
6日前
株式会社大貴
排泄物処理材及びその製造方法
今日
株式会社横島精密
太陽熱を活用した暖房システム
6日前
花王株式会社
抗菌剤組成物
6日前
井関農機株式会社
作業車両
6日前
ヤンマーホールディングス株式会社
苗移植機
今日
三州産業株式会社
蒸熱処理方法および蒸熱処理装置
10日前
花王株式会社
抗微生物組成物
10日前
オムロン株式会社
センサユニット
6日前
住友化学株式会社
複素環化合物を用いる有害節足動物防除方法
10日前
井関農機株式会社
作業車両
6日前
井関農機株式会社
作業車両
6日前
住友化学株式会社
スルホンアミド化合物を用いる有害節足動物防除方法
2日前
住友化学株式会社
スルホンアミド化合物を用いる有害節足動物防除方法
7日前
住友化学株式会社
スルホンアミド化合物を用いる有害節足動物防除方法
2日前
株式会社ブレイン
ペット用冷却マット
6日前
学校法人上智学院
ネコブセンチュウ防除剤
6日前
有限会社竹田機械
ロッドキーパー
6日前
友愉股ふん有限公司
ペット用インタラクティブ装置
9日前
井関農機株式会社
作業車両
10日前
第一金属株式会社
ラインカッター
今日
積水化学工業株式会社
魚介類の生産システムおよび魚介類の生産方法
6日前
国立大学法人 東京大学
膵炎モデル動物及びその製造方法
7日前
三井化学株式会社
抗菌性組成物
6日前
紫紘株式会社
蚕飼育システム
10日前
続きを見る
他の特許を見る




 特許ウォッチ
特許ウォッチ