TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025069175
公報種別
公開特許公報(A)
公開日
2025-04-30
出願番号
2025003800,2021523947
出願日
2025-01-09,2019-11-05
発明の名称
挿着されている細胞に直接シグナル伝達しない非天然NKG2D受容体
出願人
サイフォス、バイオサイエンシズ、インコーポレイテッド
,
XYPHOS BIOSCIENCES INC.
代理人
個人
,
個人
,
個人
,
個人
,
個人
,
個人
主分類
C07K
14/705 20060101AFI20250422BHJP(有機化学)
要約
【課題】哺乳動物細胞の表面に挿着された非天然NKG2D受容体を提供する。
【解決手段】非天然の改変NKG2D受容体であって、該受容体が、NKG2Dリガンドの非天然の改変α1-α2ドメインに結合するが、天然NKG2Dリガンドには結合せず、哺乳動物細胞に挿着された該改変NKG2D受容体が、改変α1-α2ドメインに結合している異種原子又は分子の細胞への送達のために高親和性の代替受容体として機能し、改変された該NKG2D受容体が、直接哺乳動物細胞を活性化又は哺乳動物細胞にシグナル伝達しない、受容体が提供される。
【選択図】なし
特許請求の範囲
【請求項1】
非天然の改変NKG2D受容体であって、該受容体が、NKG2Dリガンドの非天然の改変α1-α2ドメインに結合するが、天然NKG2Dリガンドには結合せず、哺乳動物細胞に挿着された該改変NKG2D受容体が、改変α1-α2ドメインに結合している異種原子又は分子の細胞への送達のために高親和性の代替受容体として機能し、改変された該NKG2D受容体が、直接哺乳動物細胞を活性化又は哺乳動物細胞にシグナル伝達しない、受容体。
続きを表示(約 1,300 文字)
【請求項2】
前記異種分子が、改変α1-α2ドメインによって哺乳動物細胞に送達される場合、哺乳動物細胞上又はその中のそれぞれの受容体サブユニットを介して該細胞を活性化又は該細胞にシグナル伝達できる、請求項1の異種分子。
【請求項3】
前記異種原子又は分子が、改変α1-α2ドメインによって哺乳動物細胞に送達される場合、非天然の改変NKG2D受容体以外の細胞上又は細胞内のいかなる受容体とも独立に、哺乳動物細胞に直接影響を及ぼし得る、請求項1の異種原子又は分子。
【請求項4】
前記異種原子又は分子が、改変α1-α2ドメインによって哺乳動物細胞に送達される場合、哺乳動物細胞によって内在化され得、改変α1-α2ドメインに結合していたか、又は非結合であったかのいずれかの、内在化された異種原子又は分子が、哺乳動物細胞に影響を与える、請求項1の異種原子又は分子。
【請求項5】
前記異種原子又は分子が、改変α1-α2ドメインによって哺乳動物細胞に送達される場合、インビボ、エクスビボ、若しくはインビトロ検出又はイメージングシステムによって検出可能なマーカーを哺乳動物細胞に提供し得る、請求項1の異種原子又は分子。
【請求項6】
共刺激ドメインが、哺乳動物細胞に導入されたエフェクター分子の機能を増強し、エフェクター分子が改変α1-α2ドメインに結合していない、前記共刺激が細胞内で結合している請求項1の非天然の改変NKG2D受容体。
【請求項7】
前記異種原子又は分子が、改変α1-α2ドメインによって哺乳動物細胞に送達される場合、補体依存性高分子攻撃複合体の形成を促進すること等の、エネルギーの放出又は細胞膜の破壊によって細胞に損傷を与え得る、請求項3の異種原子又は分子。
【請求項8】
哺乳動物細胞に挿着されている非天然の改変NKG2D受容体であって、共刺激ドメインが、細胞内で活性のあるCD3-ゼータドメインの存在なしに改変NKG2D受容体に結合しており、受容体が、NKG2Dリガンドの、非天然の改変α1-α2ドメインに結合するが、天然NKG2Dリガンドには結合しない、受容体。
【請求項9】
前記異種原子又は分子が、改変α1-α2ドメインによって哺乳動物細胞に送達される場合、哺乳動物細胞によって内在化され得、改変α1-α2ドメインに結合していたか、又は非結合であったかのいずれかの、内在化された異種原子又は分子が、哺乳動物細胞に影響を与える、異種原子又は分子が結合している請求項8の非天然の改変α1-α2ドメイン。
【請求項10】
前記異種原子又は分子が、哺乳動物細胞の、標的担持細胞又は標的担持表面への送達を、該哺乳動物細胞が標的表面又は環境に到達するときに、該哺乳動物細胞の活性化に直接つながることなく、促進するターゲティング又はホーミング機能を、該哺乳動物細胞に提供する、異種原子又は分子が結合している請求項8の非天然の改変α1-α2ドメイン。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
発明の背景
発明の分野
本出願は、大まかには、哺乳動物細胞に挿着された非天然NKG2D受容体の非天然外部ドメインであって、該非天然NKG2D受容体に特異的に結合するよう改変され、異種分子がNKG2Dリガンドの改変α1-α2ドメインに結合している非天然NKG2Dリガンドが該受容体に結合する場合、哺乳動物細胞を直接活性化又は哺乳動物細胞に直接シグナル伝達しない、外部ドメインに関する。
続きを表示(約 16,000 文字)
【背景技術】
【0002】
背景情報
NKG2Dは、ナチュラルキラー(NK)細胞及び特定のT細胞及びマクロファージの表面上にII型ホモダイマー内在性タンパク質として発現される活性化受容体である。主に、ストレスを受けている細胞の表面上で発現されているその8の天然リガンドのうちの1に結合したときに、NKG2DはNK細胞を活性化して、ストレスを受けた細胞を傷害し、または、T細胞上の場合、リガンドに占有されたNKG2Dは活性化T細胞を同時刺激して、そのエフェクター機能を実行する。ヒト天然NKG2Dの外部ドメイン、その可溶性天然リガンドのいくつか、及び、いくつかの場合、可溶性リガンド及び受容体外部ドメインの結合複合体について、三次元構造が解明されている。NKG2Dリガンドのモノマーα1-α2ドメインは、天然NKG2Dホモダイマーの2の外部ドメインに特異的に結合する。
【発明の概要】
【0003】
発明の要旨
本開示は、哺乳動物の細胞表面に挿着された非天然NKG2D受容体であって、該非天然NKG2D受容体に特異的に結合するように改変されたNKG2Dリガンドの同族非天然α1-α2ドメインが該非天然受容体に結合する場合、該受容体が該細胞に直接シグナル伝達又は該細胞を直接活性化しない、受容体に関する。NKG2Dリガンドの非天然α1-α2ドメインには、ポリペプチド、いくつかの実施形態においてはサイトカイン又は改変サイトカイン、抗体又は抗体の断片を含む異種原子又は分子が結合していてよい。細胞の直接活性化又は細胞への直接シグナル伝達は、挿着された非天然NKG2D受容体によって媒介されず、免疫学的シナプスが生じた場合でも起こらない。
図面の簡単な説明
【図面の簡単な説明】
【0004】
図1A~1B:(図1A)NKG2D.YA(配列番号18)及びNKG2D.AF(配列番号25)非天然バリアントとの天然NKG2D.wt外部ドメイン(配列番号17)のアラインメント。Y152及びY199の位置を示し、そして非天然バリアント中に存在する変異残基を灰色で強調する。(図1B)天然/野生型ULBP2の
【0005】
JPEG
2025069175000001.jpg
8
69
【0006】
及びULBP2の非天然バリアント(ULBP2.R80W(配列番号108)を含む)のアラインメント。非天然NKG2D.YAまたはNKG2D.AF受容体への非天然ULBP2バリアントの結合に重要な残基を灰色で強調する。NKG2D.YA(ULBP
2.S3、配列番号127)またはNKG2D.AF(ULBP2.C、配列番号111;ULBP2.R、配列番号113;ULPB2.AA、配列番号115;及びULBP2.AB、配列番号117)に結合する直交性バリアントについて探索したM154-F159領域及び残基R80の位置を示す。
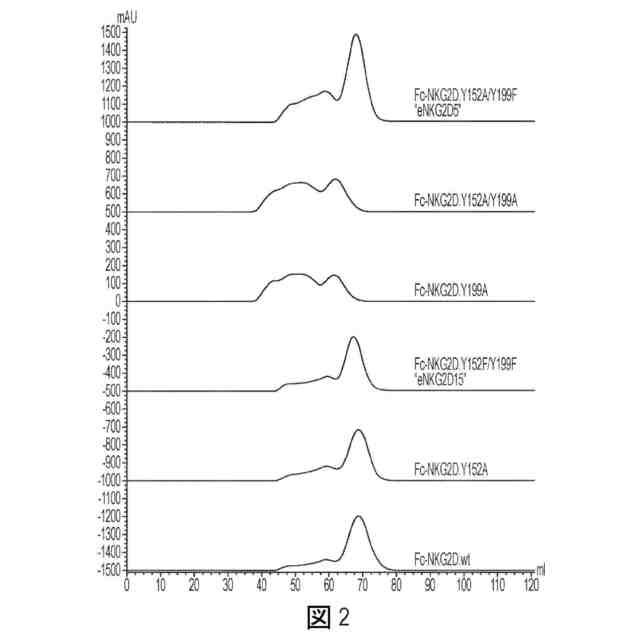
図2:Akta HiLoad 16/600 Superdex 200カラム上で分析した非天然Fc-NKG2D融合タンパク質のサイズ排除クロマトグラフィー比較。適正にアセンブルされた材料の移動は、より多い体積で溶出される分離した対称的なピークによって例示され、一方、凝集した材料はより少ない体積でより早く溶出された。改変の部位及び本質を、アミノ酸番号Y152、Y199、または両方によって示す(図の上部から開始して、配列番号48、43、42、58、41、及び40)。
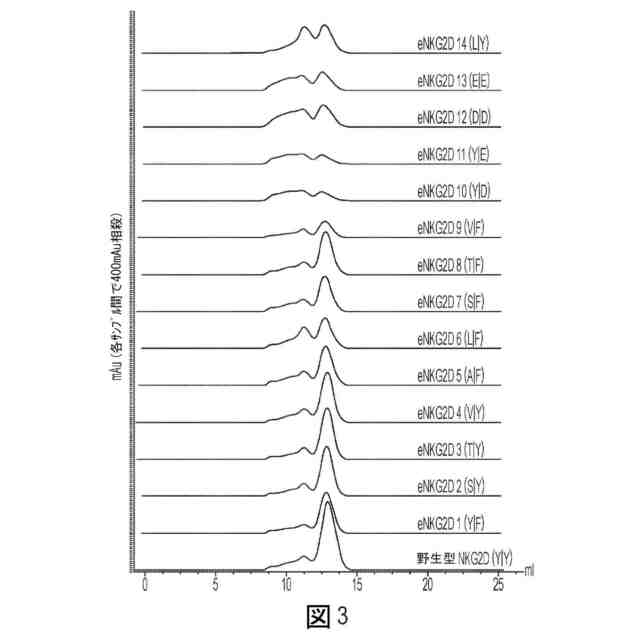
図3:1つまたは2つのアミノ酸変化を有する非天然Fc-eNKG2Dバリアントのサイズ排除クロマトグラフィープロファイルを、Akta Superdex 200 Increase 10/300 GLカラム上で分析した。適正にアセンブルされた材料の移動は、より多い体積で溶出される分離した対称的なピークによって例示され、一方、凝集した材料(低振幅のブロードなピークまたは一連のピークによって特徴付けられる)はより少ない体積で溶出された。括弧内の文字は、それぞれ152位および199位におけるアミノ酸を表す(上部から順に、配列番号57、56、55、54、53、52、51、50、49、48、47、46、45、44、および40)。
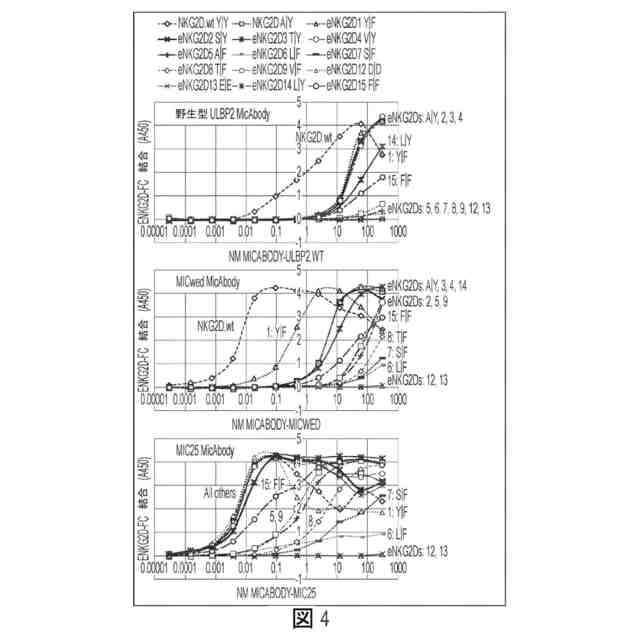
図4:Fc-eNKG2D候補へのULBP2野生型、MICwed-及びMIC25-リツキシマブMicAbodyのELISA結合。凡例を図の上部に示すが、曲線の多くは重なっているので、各々のグラフにおいて個々の曲線にラベルも付した。
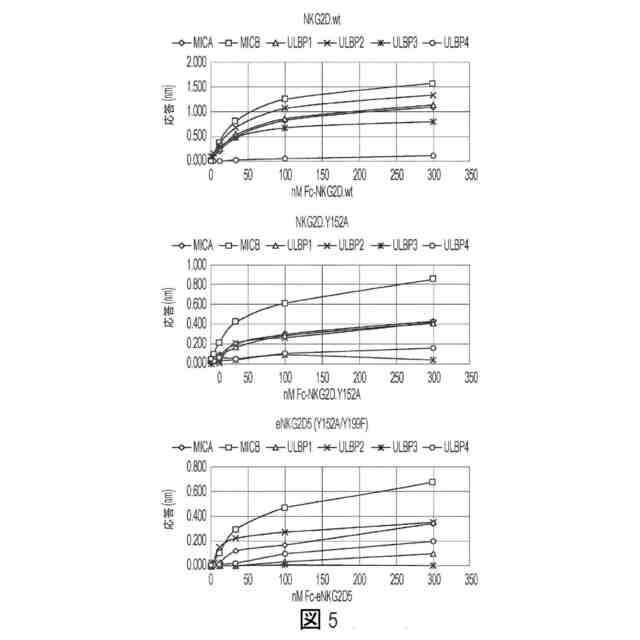
図5:野生型リガンドへのeNKG2Dバリアントの結合。野生型リガンド(全てFc融合物フォーマット)をOctet AHCバイオセンサー上に捕捉し、そしてFc融合物としての天然NKG2D、NKG2D.Y152A、またはeNKG2D5(Y152A/Y199F)の各々を300nMから0.41nMまでタイトレートした。最大結合応答をOctetによって定量した(各々グラフについて縦軸が異なることに留意のこと)。
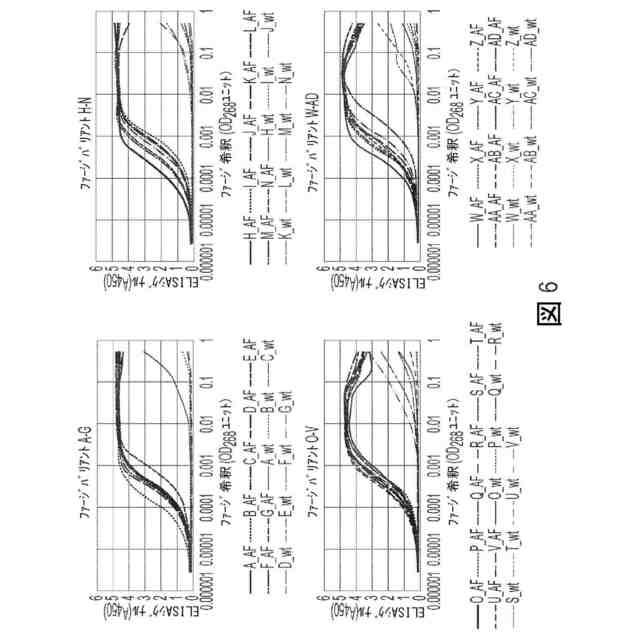
図6:Fc-NKG2D.AFへの選択的結合及びFc-NKG2D.wtへの低下または消失した結合を確認するための個別のファージバリアントのタイトレーションELISA。変異を図22において詳述する。
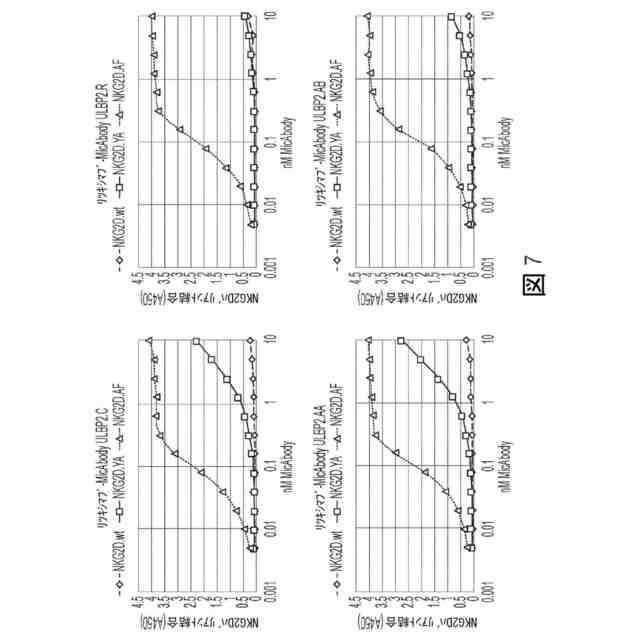
図7:NKG2D.wt、NKG2D.YA、及びNKG2D.AFに結合する4の非天然α1-α2 ULBP2バリアントMicAbodyのELISAデータ。Fc-NKG2Dバリアントを捕捉物質として使用した。MicAbodyをHRP結合抗ヒトκ中でタイトレートし、そしてそれを用いて検出した。
図8:ULBP2.C(配列番号111)、ULBP2.R(配列番号113)、ULBP2.AA(配列番号115)、及びULBP2.AB(配列番号117)を、野生型ULBP2(配列番号4)と比較してのペプチド-MHCI免疫原性の変化について、NetMHC4.0 Serverを使用し、HLAスーパータイプ代表に対してクエリーして検討した。インプット配列については、各々のバリアントについての可変領域(図1Bにおけるアラインメントに従った残基154~159)を上流及び下流の9残基とともに入れ(合計24残基)、9マーペプチドウインドウを予測免疫原性について検討した。濃灰色四角は、MHCIポケットに結合し(%順位<0.5を有するとして定義される)、それゆえ提示される強い見込みを有すると強く予測されるペプチドに対応する。淡灰色四角は、予測される弱い結合体に対応する(%順位<2)。更なる詳細については実施例5本文を参照のこと。
図8:ULBP2.C(配列番号111)、ULBP2.R(配列番号113)、ULBP2.AA(配列番号115)、及びULBP2.AB(配列番号117)を、野生型ULBP2(配列番号4)と比較してのペプチド-MHCI免疫原性の変化について、NetMHC4.0 Serverを使用し、HLAスーパータイプ代表に対してクエリーして検討した。インプット配列については、各々のバリアントについての可変領域(図1Bにおけるアラインメントに従った残基154~159)を上流及び下流の9残基とともに入れ(合計24残基)、9マーペプチドウインドウを予測免疫原性について検討した。濃灰色四角は、MHCIポケットに結合し(%順位<0.5を有するとして定義される)、それゆえ提示される強い見込みを有すると強く予測されるペプチドに対応する。淡灰色四角は、予測される弱い結合体に対応する(%順位<2)。更なる詳細については実施例5本文を参照のこと。
図9:ULBP2.C(配列番号111)、ULBP2.R(配列番号113)、ULBP2.AA(配列番号115)、及びULBP2.AB(配列番号117)を、野生型ULBP2(配列番号4)と比較してのペプチド-MHCクラスII免疫原性の変化について、NetMHCII 2.3 Serverを使用し、HLA―DR、HLA-DQ、HLA-DPに対してクエリーして検討した。インプット配列については、各々のバリアントについての可変領域(図1Bにおけるアラインメントに従った残基154~159)を上流及び下流の15残基とともに入れ(合計36残基)、15マーペプチドウインドウを予測免疫原性について検討した。濃灰色四角は、MHCIIポケットに結合し、それゆえ提示されそして免疫原性でありそうであると強く予測されるペプチドに対応する。淡灰色四角は、予測される弱い結合体に対応する。
図9:ULBP2.C(配列番号111)、ULBP2.R(配列番号113)、ULBP2.AA(配列番号115)、及びULBP2.AB(配列番号117)を、野生型ULBP2(配列番号4)と比較してのペプチド-MHCクラスII免疫原性の変化について、NetMHCII 2.3 Serverを使用し、HLA―DR、HLA-DQ、HLA-DPに対してクエリーして検討した。インプット配列については、各々のバリアントについての可変領域(図1Bにおけるアラインメントに従った残基154~159)を上流及び下流の15残基とともに入れ(合計36残基)、15マーペプチドウインドウを予測免疫原性について検討した。濃灰色四角は、MHCIIポケットに結合し、それゆえ提示されそして免疫原性でありそうであると強く予測されるペプチドに対応する。淡灰色四角は、予測される弱い結合体に対応する。
図9:ULBP2.C(配列番号111)、ULBP2.R(配列番号113)、ULBP2.AA(配列番号115)、及びULBP2.AB(配列番号117)を、野生型ULBP2(配列番号4)と比較してのペプチド-MHCクラスII免疫原性の変化について、NetMHCII 2.3 Serverを使用し、HLA―DR、HLA-DQ、HLA-DPに対してクエリーして検討した。インプット配列については、各々のバリアントについての可変領域(図1Bにおけるアラインメントに従った残基154~159)を上流及び下流の15残基とともに入れ(合計36残基)、15マーペプチドウインドウを予測免疫原性について検討した。濃灰色四角は、MHCIIポケットに結合し、それゆえ提示されそして免疫原性でありそうであると強く予測されるペプチドに対応する。淡灰色四角は、予測される弱い結合体に対応する。
図9:ULBP2.C(配列番号111)、ULBP2.R(配列番号113)、ULBP2.AA(配列番号115)、及びULBP2.AB(配列番号117)を、野生型ULBP2(配列番号4)と比較してのペプチド-MHCクラスII免疫原性の変化について、NetMHCII 2.3 Serverを使用し、HLA―DR、HLA-DQ、HLA-DPに対してクエリーして検討した。インプット配列については、各々のバリアントについての可変領域(図1Bにおけるアラインメントに従った残基154~159)を上流及び下流の15残基とともに入れ(合計36残基)、15マーペプチドウインドウを予測免疫原性について検討した。濃灰色四角は、MHCIIポケットに結合し、それゆえ提示されそして免疫原性でありそうであると強く予測されるペプチドに対応する。淡灰色四角は、予測される弱い結合体に対応する。
図9:ULBP2.C(配列番号111)、ULBP2.R(配列番号113)、ULBP2.AA(配列番号115)、及びULBP2.AB(配列番号117)を、野生型ULBP2(配列番号4)と比較してのペプチド-MHCクラスII免疫原性の変化について、NetMHCII 2.3 Serverを使用し、HLA―DR、HLA-DQ、HLA-DPに対してクエリーして検討した。インプット配列については、各々のバリアントについての可変領域(図1Bにおけるアラインメントに従った残基154~159)を上流及び下流の15残基とともに入れ(合計36残基)、15マーペプチドウインドウを予測免疫原性について検討した。濃灰色四角は、MHCIIポケットに結合し、それゆえ提示されそして免疫原性でありそうであると強く予測されるペプチドに対応する。淡灰色四角は、予測される弱い結合体に対応する。
図9:ULBP2.C(配列番号111)、ULBP2.R(配列番号113)、ULBP2.AA(配列番号115)、及びULBP2.AB(配列番号117)を、野生型ULBP2(配列番号4)と比較してのペプチド-MHCクラスII免疫原性の変化について、NetMHCII 2.3 Serverを使用し、HLA―DR、HLA-DQ、HLA-DPに対してクエリーして検討した。インプット配列については、各々のバリアントについての可変領域(図1Bにおけるアラインメントに従った残基154~159)を上流及び下流の15残基とともに入れ(合計36残基)、15マーペプチドウインドウを予測免疫原性について検討した。濃灰色四角は、MHCIIポケットに結合し、それゆえ提示されそして免疫原性でありそうであると強く予測されるペプチドに対応する。淡灰色四角は、予測される弱い結合体に対応する。
図9:ULBP2.C(配列番号111)、ULBP2.R(配列番号113)、ULBP2.AA(配列番号115)、及びULBP2.AB(配列番号117)を、野生型ULBP2(配列番号4)と比較してのペプチド-MHCクラスII免疫原性の変化について、NetMHCII 2.3 Serverを使用し、HLA―DR、HLA-DQ、HLA-DPに対してクエリーして検討した。インプット配列については、各々のバリアントについての可変領域(図1Bにおけるアラインメントに従った残基154~159)を上流及び下流の15残基とともに入れ(合計36残基)、15マーペプチドウインドウを予測免疫原性について検討した。濃灰色四角は、MHCIIポケットに結合し、それゆえ提示されそして免疫原性でありそうであると強く予測されるペプチドに対応する。淡灰色四角は、予測される弱い結合体に対応する。
図9:ULBP2.C(配列番号111)、ULBP2.R(配列番号113)、ULBP2.AA(配列番号115)、及びULBP2.AB(配列番号117)を、野生型ULBP2(配列番号4)と比較してのペプチド-MHCクラスII免疫原性の変化について、NetMHCII 2.3 Serverを使用し、HLA―DR、HLA-DQ、HLA-DPに対してクエリーして検討した。インプット配列については、各々のバリアントについての可変領域(図1Bにおけるアラインメントに従った残基154~159)を上流及び下流の15残基とともに入れ(合計36残基)、15マーペプチドウインドウを予測免疫原性について検討した。濃灰色四角は、MHCIIポケットに結合し、それゆえ提示されそして免疫原性でありそうであると強く予測されるペプチドに対応する。淡灰色四角は、予測される弱い結合体に対応する。
図9:ULBP2.C(配列番号111)、ULBP2.R(配列番号113)、ULBP2.AA(配列番号115)、及びULBP2.AB(配列番号117)を、野生型ULBP2(配列番号4)と比較してのペプチド-MHCクラスII免疫原性の変化について、NetMHCII 2.3 Serverを使用し、HLA―DR、HLA-DQ、HLA-DPに対してクエリーして検討した。インプット配列については、各々のバリアントについての可変領域(図1Bにおけるアラインメントに従った残基154~159)を上流及び下流の15残基とともに入れ(合計36残基)、15マーペプチドウインドウを予測免疫原性について検討した。濃灰色四角は、MHCIIポケットに結合し、それゆえ提示されそして免疫原性でありそうであると強く予測されるペプチドに対応する。淡灰色四角は、予測される弱い結合体に対応する。
図10A~10B:天然NKG2D.wt、NKG2D.YA、及びNKG2D.AFへの、ULBP2.wt(野生型)、ULBP2.R80W(野生型NKG2Dに対する増強された親和性を有する)、ULPB2.S3(NKG2D.YA選択された直交性バリアント)、またはULBP2.R(NKG2D.AF選択された直交性バリアント)を含むリツキシマブ-MicAbodyのELISA測定された結合。(図10A)ELISA曲線。より高い濃度でのいくつかのアッセイについての458nm吸収の低下は、TMB-Ultra ELISA発色試薬の沈殿に起因して、より高い濃度の高親和性係合物(engager)についてしばしば見られるアーチファクトである。(図10B)(A)における曲線に基づいてGraphPad Prismにおいて決定されたEC50値(nMで報告する)。nd=増大した濃度と結合との間の関係の欠如に起因して決定されず。
図11A~11C:形質導入されていないかまたは、CD8aヒンジ/膜貫通ドメイン並びに細胞内4-1BB及びCD3ゼータシグナル伝達ドメインからなるNKG2D.wt、NKG2D.YA、若しくはNKG2D.AF CAR構築物を用いて形質導入されたかのいずれかのCD8エフェクター細胞を用いたインビトロ細胞溶解アッセイ。標的細胞にカルセインを前負荷し、そして漸増するエフェクター対標的(E:T)比でエフェクター細胞に曝露した。放出されたカルセインを5時間後に定量した。(図11A)HeLa細胞の細胞溶解。(図11B)表面ULBP1を過剰発現するようにトランスフェクトされたHeLa細胞の溶解。(図11C)その表面上に非天然ULBP2.Rを発現するHeLa細胞の細胞溶解。エラーバーは、実験における技術的複製物の標準偏差に対応する。
図12A~12B:NKG2D-CAR CD8 T細胞による腫瘍株のMicAbody指図細胞溶解。(図12A)Ramos細胞(リツキシマブの標的であるCD20を発現する)にカルセインを前負荷し、そして漸増濃度のULBP2.S2またはULBP2.Rリツキシマブ-Micabodyのいずれかとともに20:1のE:T比でNKG2D.AF-またはNKG2D.YA-CAR細胞に曝露した。2時間の共インキュベーションの後に細胞溶解のレベルを定量した。(図12B)ヒトHer2を発現するようにトランスフェクトしたマウス腫瘍株CT26を、Ramos細胞と並行して細胞溶解標的として使用した。NKG2D.AF-CAR CD8 T細胞に飽和濃度(5nM)のリツキシマブ-ULBP2.R、トラスツズマブ-ULBP2.R、または2のMicAbodyの等モル混合物を前付与した。非結合MicAbodyを洗浄によって除去し、そしてCD8細胞を標的細胞に2の異なるE:T比で添加した。2時間後に細胞溶解を測定した。
図13A~13D:MicAbody及びMicAdaptorフォーマットの候補の説明。(図13A)MicAbody及びMicAdaptor試薬の開発において利用されるさまざまな抗体Fcバリアントには、(a)野生型ヒトIgG1 Fc、(b)FcをADCC欠損にする2の変異、及び(c)ADCC欠損変異も含有するヘテロ二量体Fc分子の生成を可能にする、それぞれのFc―Fc1又はFc2―における静電ステアリング変異(electrostatic steering mutation)が含まれる。(図13B)直交性リガンドを(a)重鎖又は(b)軽鎖のC末端に融合させてMicAbody試薬を生成する方法の例。(図13C)抗体成分を含まない直接の直交性リガンド融合物を伴うMicAdaptorの例。(図13D)血清安定性を増強するためにヒトIgG1 Fcとの関連で生成され得るさまざまなMicAdaptor分子の説明であり、異種カーゴ又は直交性リガンドのいずれかについての所望の分子の価数に応じて、ヘテロ二量体-Fc変異を含む(a、b、c、本文及び図の説明では「Fc1/Fc2」と表記)か、又は含まない(d、e)場合がある。これは、所望のカーゴ、価数、及び機能性(f、g)によっては完全な抗体構造を利用することも含み得、重鎖又は軽鎖のいずれかの融合物であり得る。
図14:CAR構築物、サイレントCAR、及び他のCARバリアントの模式図。各構築物についての配列番号を表示。
図15A~15C:NKG2D.YA-CARを発現するCD8 T細胞へのサイトカイン融合物の選択的送達。(図15A)NKG2D.wt-CAR(配列番号151)又はNKG2D.YA-CAR 配列番号153)のいずれかを発現するCD8細胞を30又は300IUe/mLの組換えヒトIL2(rhIL2)又はIL15(rhIL15)、又は変異型-IL2/-IL15直交性リガンドの直接の融合物又はヘテロ二量体Fc(Fc1/Fc2)との関連での融合物の変形に曝露した。3日後、WST細胞増殖試薬により増殖を定量化した。ここでのデータは、サイトカインなしのコントロールに対して正規化して示す。(図15B)NKG2D.YA-CARをコードするベクターで形質導入したヒトT細胞を様々なリガンド-サイトカイン融合物分子に7日間曝露し、培養物中のGFP+CAR-細胞のパーセンテージを経時的に追跡した。(図15C)直交性リガンドは、NKG2D.YA-CAR細胞へのIL21及び変異体IL21試薬の送達を増強し、WSTアッセイによって決定されるように、非形質導入細胞と比較して、3日間の培養にわたって、それらの増幅を促進する。サイトカイン又はサイトカイン-MicAdaptorのIUe/mLを、横軸の説明中の括弧内に示す。MicAdaptorの配列番号については、図24を参照。
図16A~16C:細胞への直交性リガンド-サイトカイン-融合物の送達の促進におけるNKG2D.YA外部ドメイン単独の十分性を調査するデータ。(図16A)NKG2D.YA外部ドメイン単独(配列番号157)は、リツキシマブ-ULBP2.S3 MicAbody(配列番号98及び129)によるRamos細胞の殺傷を指図することができなかった。(図16B)様々なサイトカイン試薬への3日間の曝露後のWST増殖アッセイ。(図16C)WST増殖アッセイにより、NKG2D.YA-CARの直交性リガンドとの係合(engagement)は、細胞増殖を促進するのには不十分(但し、融合サイトカイン又はサイトカイン変異体の非存在下で)であることが実証された。MicAdaptorの配列番号については、図24を参照。サイトカイン及びサイトカイン-MicAdaptorのIUE/mL量を、横軸の説明中の括弧内に示す。
図17A~17C:(図17A)示したサイトカイン又はサイトカイン-MicAdaptorとの3日間のインキュベーション後の、様々な共刺激ドメイン変異体(配列番号161、163、及び165)のWST増殖アッセイ。(図17B)リツキシマブ-ULBP2.S3 MicAbod(配列番号98及び129)の存在下で、カルセインをロードしたRamos標的細胞を効果的に溶解するそれらの能力について調べた同一の共刺激ドメインCAR変異体。(図17C)サイトカイン試薬との3日間のインキュベーション後のWSTアッセイによって評価された通り、NKG2D.YA外部ドメイン(NKG2D.YA-ecd)と完全なCD19scFv-CARとの共発現は、増殖を促進するのに十分である。MicAdaptorの配列番号については、図24を参照。
図18:サイズ排除クロマトグラフィー(SEC)によって決定された、候補の非天然Fc-eNKG2Dバリアントの変異及びタンパク質凝集特性のまとめ。
図19:MICwed-MicAbodyまたはMIC25-MicAbodyのいずれかによる野生型NKG2D結合に対して正規化されたeNKG2Dバリアントの飽和百分率(Rmax)。野生型Fc-NKG2D及び各々のFc-eNKG2D受容体をAHCバイオセンサー上に捕捉し、次いで20nMのトラスツズマブ特異的MicAbodyに曝露した。解離速度をモニターし、そしてFc-eNKG2D融合物のRmax値を順位付けした。試験しなかったサンプル(nt)は、激しい凝集、または発現されたか若しくはSEC分画後に回収された材料の不十分な量のいずかに起因した。
図20:図3に示すFc-eNKG2D ELISAについてのEC50値(nM)。nt=試験せず;nb=結合なし、または300nMでさえ非常に低い結合で、EC50値算出されず。
図21:スポットELISAによって確認した、天然NKG2D.wtに対比してNKG2D.AFへの選択的結合を有するファージクローンをもたらしたULBP2内の組み合わせ変異のサブセット。変異を、選択されたファージの中での出現頻度によって順位付けした。
図21:スポットELISAによって確認した、天然NKG2D.wtに対比してNKG2D.AFへの選択的結合を有するファージクローンをもたらしたULBP2内の組み合わせ変異のサブセット。変異を、選択されたファージの中での出現頻度によって順位付けした。
図22:リツキシマブ-MicAbodyフォーマットのNKG2D.AF選択されたULBP2バリアントの特異性は、定量的ELISAにより、NKG2D.AFへのその結合を保持した。各々のULBP2バリアントの特異的アミノ酸改変を、Fc-NKG2D.AF融合物に対比したFc-NKG2D.wt融合物へのその結合の比として示す。ULBP2のアミノ酸残基の位置は図1Bのものである。
図23:Y152A特異的ファージクローンをもたらした、ULBP2.R80W(図1B;配列番号108)の表示するアミノ酸位置における、選択された変異。
図24:MicAdaptorの配列番号及び一過性トランスフェクションからの精製方法。
【発明を実施するための形態】
【0007】
発明の詳細な説明
免疫系のナチュラルキラー(NK)細胞及び特定の(CD8+αβ及びγδ)T細胞は、ヒト及び他の哺乳動物において、新生物及び感染細胞に対する第一線の生得的な防御として重要な役割を有している(Cerwenka,A.,and L.L.Lanier.2001.NK cells,viruses and cancer.Nat.Rev.Immunol.1:41-49)。NK細胞及び特定のT細胞は、標的細胞の認識及び病態細胞に対する生得的な防御の活性化を担う顕著なホモダイマー表面免疫受容体であるNKG2Dをその表面上に示す(Lanier,LL,1998.NK cell receptors.Ann.Rev.Immunol.16:359-393;Houchins JP et al.1991.DNA sequence analysis of NKG2,a family of related cDNA clones encoding type II integral membrane proteins on human NK cells.J.Exp.Med.173:1017-1020;Bauer,S et al.,1999.Activation of NK cells and T cells by NKG2D,a receptor for stress-inducible MICA.Science 285:727-730)。ヒトNKG2D分子は、その8の異なる同族リガンドに結合するC型レクチン様細胞外(外部)ドメインを有しており、最も研究されているリガンドは84%配列同一または相同のモノマーMICA及びMICB(主要組織適合性複合体(MHC)クラスI鎖関連糖タンパク質(MIC)の多型アナログ)である(Weis et al.1998.The C-type lectin superfamily of the immune system.Immunol.Rev.163:19-34;Bahram et al.1994.A second lineage of mammalian MHC class I genes.PNAS 91:6259-6263;Bahram et al.1996a.Nucleotide sequence of the human MHC class I MICA gene.Immunogenetics 44: 80-81;Bahram and Spies TA.1996.Nucleotide sequence of human MHC
class I MICB cDNA.Immunogenetics 43:230-233)。MICA及びMICBの非病態的発現は、一部の腸上皮、ケラチノサイト、内皮細胞及び単球に制限されているが、これらのMICタンパク質の異常な表面発現が、増殖、酸化及び熱ショックのような多くの型の細胞ストレスに応答して生じ、そして細胞を病態的と特徴付ける(Groh et al.1996.Cell stress-regulated human MHC class I gene expressed in GI epithelium.PNAS 93:12445-12450;Groh et al.1998.Recognition of stress-ind
uced MHC molecules by intestinal γδ T cells.Science 279:1737-1740;Zwirner et al.1999.Differential expression of MICA by endothelial cells,fibroblasts,keratinocytes and monocytes.Human Immunol.60:323-330)。MICタンパク質の病態的発現はまた、いくつかの自己免疫疾患に関与しているようである(Ravetch,JV and Lanier LL.2000.Immune Inhibitory Receptors.Science 290:84-89;Burgess,SJ.2008.Immunol.Res.40:18-34)。多型MICA及びMICBのようなNKG2Dリガンドの差次的調節は、所望されない攻撃から健常細胞を依然として防御しながら広範な緊急の合図を同定しそれに応答するための手段を免疫系に提供するために重要である(Stephens HA,(2001)MICA and MICB genes:can the enigma of their polymorphism be resolved? Trends Immunol.22:378-85;Spies,T.2008.Regulation of
NKG2D ligands: a purposeful but delicate affair.Nature Immunol.9:1013-1015)。
【0008】
ウイルス感染は、MICタンパク質発現の一般的な誘導因子であり、NKまたはT細胞の攻撃のためにウイルス感染細胞を同定する(Groh et al.1998;Groh et al.2001.Co-stimulation of CD8+ αβT cells by NKG2D via engagement by MIC induced on virus-infected cells.Nat.Immunol.2:255-260;Cerwenka,A.,and L.L.Lanier.2001)。事実、その宿主細胞上でのそのような攻撃を回避するために、サイトメガロウイルス及び他のウイルスは、自然免疫系による標的化から逃避するために、それが感染する細胞の表面上でのMICタンパク質の発現を防止する進化した機構を有する(Lodoen,M.,K.Ogasawara,J.A.Hamerman,H.Arase,J.P.Houchins,E.S.Mocarski,and L.L.Lanier.2003.NKG2D-mediated NK cell protection against cytomegalovirus is impaired by gp40 modulation of RAE-1 molecules.J.Exp.Med.197:1245-1253;Stern-Ginossar et al.,(2007)Host immune system gene targeting by
viral miRNA.Science 317:376-381;Stern-Ginossar et al.,(2008)Human microRNAs regulate stress-induced immune responses mediated by the receptor NKG2D.Nature Immunology 9:1065-73;Slavuljica,I A Busche,M
Babic,M Mitrovic,I Gasparovic,D Cekinovic,E Markova Car,EP Pugel,A Cikovic,VJ Lisnic,WJ Britt,U Koszinowski,M Messerle,A Krmpotic and S Jonjic.2010.Recombinant
mouse cytomegalovirus expressing a ligand for the NKG2D receptor is attenuated and has improved vaccine properties.J.Clin.Invest.120:4532-4545)。
【0009】
それらのストレスにもかかわらず、肺がんや神経膠芽細胞腫脳がんのもの等の多くの悪性細胞も、MICタンパク質の発現を回避し、その結果、自然免疫系から過剰に逃れるた
め、特に攻撃的である可能性がある(Busche,A et al.2006,NK cell mediated rejection of experimental human lung cancer by genetic over expression of MHC class I chain-related gene A.Human Gene Therapy 17:135-146;Doubrovina,ES,MM Doubrovin,E Vider,RB Sisson,RJ O’Reilly,B Dupont,and YM Vyas,2003.Evasion from NK Cell Immunity by MHC Class I Chain-Related Molecules Expressing Colon Adenocarcinoma (2003)J.Immunology 6891-99;Friese,M.et al.2003.MICA/NKG2D-mediated immunogene therapy of experimental gliomas.Cancer Research 63:8996-9006;Fuertes,MB,MV Girart,LL Molinero,CI Domaica,LE
Rossi,MM Barrio,J Mordoh,GA Rabinovich and NW Zwirner.(2008)Intracellular Retention of the NKG2D Ligand MHC Class I Chain-Related Gene A in Human Melanomas Confers Immune Privilege and Prevents NK Cell-Mediated Cytotoxicity.J.Immunology,180:4606-4614)。
【0010】
NKG2Dに結合したヒトMICAの高分解能構造が解明されており、それによってMICAのα3ドメインがNKG2Dとの直接的な相互作用を有しないことが実証されている(Li et al. 2001. Complex structure of the activating immunoreceptor NKG2D andits MHC class I-like ligand MICA. Nature Immunol. 2: 443-451; Protein Data Bankaccession code 1HYR)。MICAのα3ドメインは、MICBのもののように、短い可動性のリンカーペプチドによってα1-α2プラットフォームドメインに連結されており、そしてそれ自体はプラットフォームとMIC発現細胞の表面との間の「スペーサー」として天然に配置されている。ヒトMICA及びMICBα3ドメインの三次元構造はほぼ同一であり(94C-αα’の二乗平均平方根距離<1Å)、そして機能的に互換的である(Holmes et al.2001.Structural Studies of Allelic Diversity of the MHC Class I Homolog MICB, a Stress-Inducible Ligand for the Activating Immunoreceptor NKG2D.J Immunol.169:1395-1400)。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
東ソー株式会社
炭素-窒素結合形成方法
11日前
株式会社トクヤマ
四塩化炭素の製造方法
11日前
株式会社トクヤマ
シロキサン類の回収方法
11日前
日産化学株式会社
ピリジン化合物の製造方法
2か月前
日本特殊陶業株式会社
メタン製造装置
1か月前
信越化学工業株式会社
新規化合物
13日前
金剛化学株式会社
ボルチオキセチンの製造方法
1か月前
artience株式会社
四塩基酸無水物の製造方法
18日前
ユニチカ株式会社
ビスマレイミドおよびその製造方法
1か月前
四国化成工業株式会社
エポキシ化合物およびその利用
2か月前
国立大学法人京都大学
細胞質送達ペプチド
3か月前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
21日前
四国化成工業株式会社
イソフタル酸化合物およびその利用
2か月前
株式会社トクヤマ
サフィナミド若しくはその塩の製造方法
18日前
四国化成工業株式会社
テレフタル酸化合物およびその利用
2か月前
キヤノン株式会社
有機化合物及び有機発光素子
2か月前
キヤノン株式会社
有機化合物及び有機発光素子
1か月前
小川香料株式会社
化合物及び香料組成物
3か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の保存溶液
18日前
キヤノン株式会社
有機化合物及び有機発光素子
1か月前
株式会社カネカ
プロピレンオキサイド(PO)製造システム
2か月前
旭化成株式会社
トリオキサンの製造方法
21日前
小川香料株式会社
化合物及び香料組成物
3か月前
ダイキン工業株式会社
SF5含有シラン化合物
3か月前
キヤノン株式会社
有機化合物及び有機発光素子
1か月前
株式会社半導体エネルギー研究所
有機金属錯体、発光デバイス
1か月前
JFEスチール株式会社
メタノール合成方法
1か月前
旭化成株式会社
ホルムアルデヒドの製造方法
1か月前
公立大学法人大阪
カルボン酸化合物の製造方法
14日前
株式会社半導体エネルギー研究所
有機化合物、および発光デバイス
25日前
株式会社トクヤマ
精製第4級アンモニウム化合物水溶液の製造方法
2か月前
出光興産株式会社
プロピレンの製造方法
1か月前
旭化成株式会社
軽質オレフィンの製造方法
1か月前
住友ベークライト株式会社
フェノールの製造方法
1か月前
JX金属株式会社
高純度ギ酸銅
13日前
日産化学株式会社
新規な縮合ヘテロ環アミド化合物及び有害生物防除剤
1か月前
続きを見る
他の特許を見る




 特許ウォッチ
特許ウォッチ