TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025012406
公報種別
公開特許公報(A)
公開日
2025-01-24
出願番号
2023115213
出願日
2023-07-13
発明の名称
イソアスパラギン酸(Lβ-アスパラギン酸)残基を標識する方法
出願人
国立研究開発法人産業技術総合研究所
代理人
個人
,
個人
,
個人
,
個人
主分類
G01N
27/62 20210101AFI20250117BHJP(測定;試験)
要約
【課題】本発明は、アスパラギン酸残基を含む対象ペプチド中のLβ-アスパラギン酸残基を標識する方法に関する。
【解決手段】本発明の方法は、(1)ペプチド中のLα-アスパラギン酸残基を特異的に認識してペプチドを切断するプロテアーゼで、対象ペプチドを処理することによって、Lα-アスパラギン酸残基を有するペプチドを分解する工程、(2)反応系から前記Lα-アスパラギン酸残基を特異的に認識してペプチドを切断するプロテアーゼを除去する工程、(3)Lβ-アスパラギン酸残基を特異的にメチルエステル化し、Lα-アスパラギン酸残基へ変換するメチルトラスフェラーゼにより、Lβ-アスパラギン酸残基をメチルエステル化する工程、ここにおいて、工程(3)を
18
O水中で行うことにより、変換後のLα-アスパラギン酸残基のヒドロキシ基が
18
Oで標識される、を含む。
【選択図】なし
特許請求の範囲
【請求項1】
アスパラギン酸残基を含む対象ペプチド中のLβ-アスパラギン酸残基を標識する方法であって、
(1)ペプチド中のLα-アスパラギン酸残基を特異的に認識してペプチドを切断するプロテアーゼで、対象ペプチドを処理することによって、Lα-アスパラギン酸残基を有するペプチドを分解する工程、
(2)反応系から前記Lα-アスパラギン酸残基を特異的に認識してペプチドを切断するプロテアーゼを除去する工程、
および
(3)Lβ-アスパラギン酸残基を特異的にメチルエステル化し、Lα-アスパラギン酸残基へ変換するメチルトラスフェラーゼにより、Lβ-アスパラギン酸残基をメチルエステル化する工程、
ここにおいて、工程(3)を
18
O水中で行うことにより、変換後のLα-アスパラギン酸残基のヒドロキシ基が
18
Oで標識される、
を含む、前記方法。
続きを表示(約 1,500 文字)
【請求項2】
工程(3)のLβ-アスパラギン酸残基を特異的にメチルエステル化し、Lα-アスパラギン酸残基へ変換するメチルトラスフェラーゼが、L-イソアスパラギン酸-メチルトランスフェラーゼである、請求項1に記載の方法。
【請求項3】
前記対象ペプチドが、3-100アミノ酸残基の長さのペプチドである、請求項1に記載の方法。
【請求項4】
前記対象ペプチドが、タンパク質を分解するプロテアーゼによって、タンパク質を処理することにより得たものである、請求項1に記載の方法。
【請求項5】
工程(1)の前に、
(0-i)タンパク質を分解するプロテアーゼによって、対象タンパク質を処理し、対象ペプチドを得る工程、
(0-ii)反応系から前記タンパク質を分解するプロテアーゼを除去する工程
を含む、請求項1に記載の方法。
【請求項6】
工程(0-i)のタンパク質を分解するプロテアーゼによる処理工程を、pH8~9の条件下で行う、請求項5に記載の方法。
【請求項7】
前記タンパク質を分解するプロテアーゼが、トリプシン、キモトリプシン、エラスターゼ、テルモリシン、ペプシン、Glu-C、Glu-N、Lys-C、Lys-N、Arg-C、V8プロテアーゼ、IdeS、IdeZ、およびコラゲナーゼからなる群から選択される、請求項4または5に記載の方法。
【請求項8】
(1)ペプチド中のLα-アスパラギン酸残基を特異的に認識してペプチドを切断するプロテアーゼで、対象ペプチドを処理することによって、Lα-アスパラギン酸残基を有するペプチドを分解する工程、または、(0-i)タンパク質を分解するプロテアーゼによって、対象タンパク質を処理し、対象ペプチドを得る工程、から、(3)Lβ-アスパラギン酸残基を特異的にメチルエステル化し、Lα-アスパラギン酸残基へ変換するメチルトラスフェラーゼにより、Lβ-アスパラギン酸残基をメチルエステル化する工程、まで同一pHのバッファーを使用する、請求項1-5のいずれか1項に記載の方法。
【請求項9】
対象タンパク質を分析する方法であって、
(0-i)タンパク質を分解するプロテアーゼによって、対象タンパク質を処理し、対象ペプチドを得る工程、
(0-ii)反応系から前記タンパク質を分解するプロテアーゼを除去する工程、
(1)ペプチド中のLα-アスパラギン酸残基を特異的に認識してペプチドを切断するプロテアーゼで、前記対象ペプチドを処理することによって、Lα-アスパラギン酸残基を有するペプチドを分解する工程、
(2)反応系から前記Lα-アスパラギン酸残基を特異的に認識してペプチドを切断するプロテアーゼを除去する工程、
(3)Lβ-アスパラギン酸残基を特異的にメチルエステル化し、Lα-アスパラギン酸残基へ変換するメチルトラスフェラーゼにより、Lβ-アスパラギン酸残基をメチルエステル化する工程、
ここにおいて、工程(3)を
18
O水中で行うことにより、変換後のLα-アスパラギン酸残基のヒドロキシ基が
18
Oで標識される、
および、
(4)工程(3)で標識されたペプチド中のアスパラギン酸残基を質量分析する工程、
を含む、前記方法。
【請求項10】
工程(4)の質量分析は、LC-MSまたはMALDI-MSによって行う、請求項9に記載の方法。
発明の詳細な説明
【技術分野】
【0001】
本発明は、アスパラギン酸残基を含む対象ペプチド中のイソアスパラギン酸(Lβ-アスパラギン酸)残基を標識する方法に関する。本発明はまた、対象タンパク質中のLβ-アスパラギン酸残基を分析する方法に関する。
続きを表示(約 1,800 文字)
【背景技術】
【0002】
アミノ酸にはL型とD型の二つの鏡像異性体がある。長い間、タンパク質を構成するアミノ酸は全てL-アミノ酸であると考えられてきた。しかし、近年、D-アミノ酸を含むタンパク質やペプチドが生体の種々の組織で発見されてきている。
【0003】
アミノ酸残基の異性化は、タンパク質の立体構造を変化させ、タンパク質機能に影響を与えることが示唆されている(非特許文献1-3)。従って、アミノ酸異性体を含むタンパク質の蓄積は、種々の疾患の発症に関連していると考えられる。
【0004】
特に、アスパラギン酸(Asp)残基は、タンパク質を構成するアミノ酸の中で最も異性化しやすいアミノ酸である。非特許文献4は、合成ペプチドを用いて各Asp異性体への異性化速度を計算し、ライフスパンの間のAsp異性体の蓄積をシミュレートしたものを記載している。タンパク質代謝がなければ、Lα-Aspは数年で急速にLβ-Aspに置き換わり、その後Dα-AspとDβ-Aspに徐々に置き換わることが報告されている(非特許文献2)。この結果は、Lβ-Aspの大規模同定のための技術がAsp異性化タンパク質の包括的探索において非常に重要であることを示す。
【0005】
Aspは、特有の異性化機構を有しており、生体の温和な条件下で異性化が進行し、通常のLα体からLβ、Dα、Dβの3つの異性体ヘと異性化する。Aspは、以下の式1に示すように、スクシンイミドによって媒介される化学平衡反応のために、Asp残基のα水素を直接引抜くことなく異性化される。
【0006】
JPEG
2025012406000001.jpg
86
152
【0007】
Aspの異性化は化学平衡反応により生じることから、生体内における異性化の主たる要因は時間であると考えられている。したがって、Asp残基の異性化は、水晶体や骨などの代謝回転の遅い組織中のタンパク質で起こると考えられている。特に水晶体では、胎児期に形成されたタンパク質が生涯代謝されないため、異性化タンパク質が蓄積している。さらに、白内障患者の水晶体タンパク質のAsp異性化率は同年齢の健常者よりも高く、Asp異性化は白内障発症の引き金であると考えられている(非特許文献5)。このように、Aspの異性化は、これまでターンオーバーの長い限られた組織においてのみ蓄積・検出できるものと考えらえてきた。しかしながら、近年、血清中のIgGなどにおいても、Asp異性体の存在が確認されており、Asp異性体が生体内に広く存在する可能性が示唆されている(非特許文献1)。
【0008】
また、タンパク質中およびペプチド中のアスパラギン(Asn)残基は脱アミド化して、アスパラギン酸残基と同一の構造であるL-スクシンイミド体を形成し、アスパラギン酸残基へと変換される。すなわち、アスパラギン酸異性体はAsp残基およびAsn残基から形成される。
【0009】
質量分析によるプロテオミクスは、タンパク質に起こる翻訳後修飾の大規模な探索に有用である。しかしながら、Asp異性化は、分子量の変化を伴わないので、リン酸化やグリコシル化のようにプロテオミクスへ適用させることは難しい。
【0010】
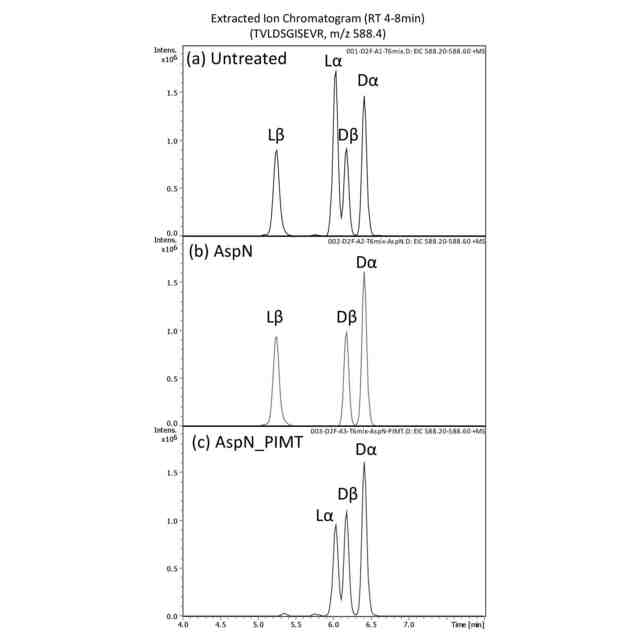
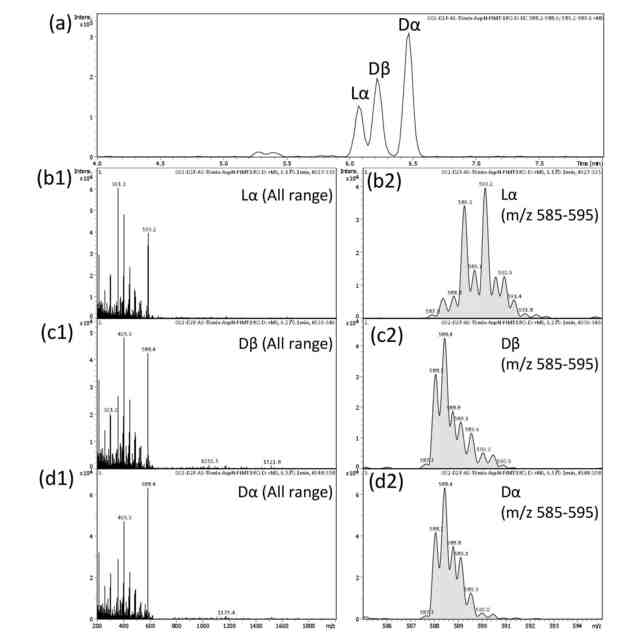
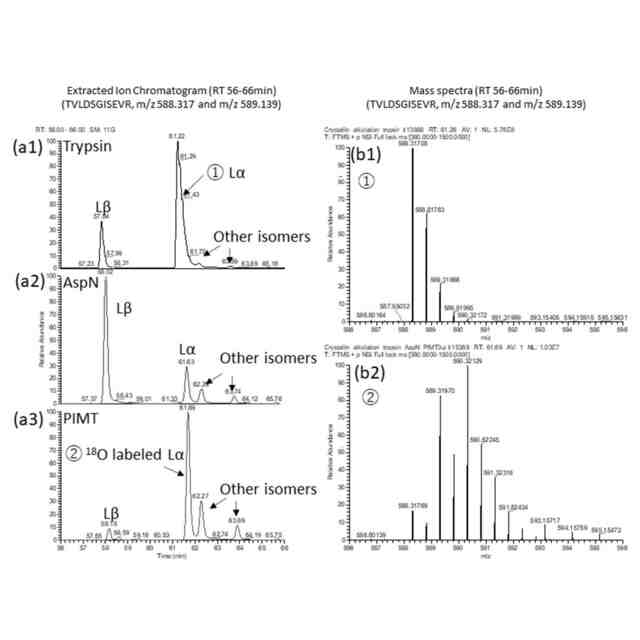
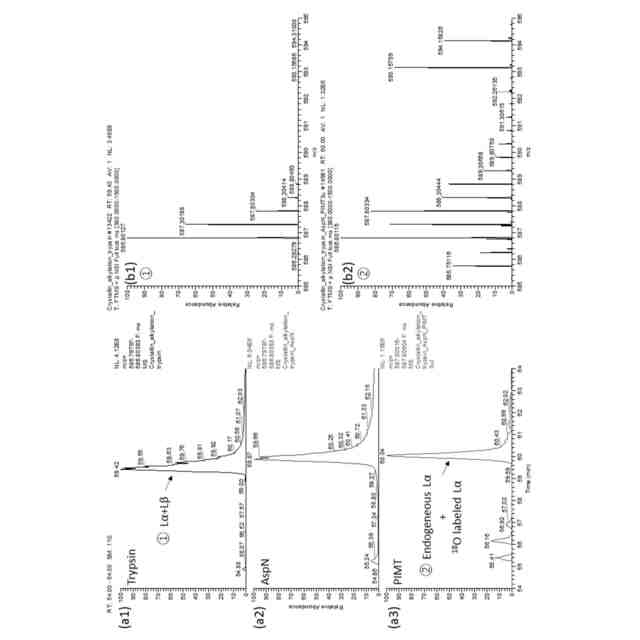
本発明者らは以前、LCクロマトグラムにおいて、Asp異性体を有するペプチドが異なる溶出時間で溶出するという特徴に基づいて、Asp異性体ペプチドを同定する方法を報告した(非特許文献6、7)。これは、溶出時間を合成ペプチドと一致させるか、あるいは、Asp異性体を特異的に切断または変換する酵素を用いることによって達成することができる。具体的には、Lα-Aspは、エンドペプチダーゼ「AspN」によって切断し、Lβ-Aspには、プロテイン D-アスパラギン酸 メチルトランスフェラーゼ(protein D-aspartate methyltransferase:PIMT)を作用させ、そして、Dα-Aspには、D-アスパラギン酸エンドペプチダーゼ(DAEP)で分解する、というものである。しかし、この方法では分析されるペプチドを1つずつ選択する必要があり、Asp異性体ペプチドを大規模に探索することは困難である。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
個人
集束超音波の測定機
18日前
個人
センサ制御回路
18日前
甲神電機株式会社
電流検出器
11日前
甲神電機株式会社
漏電検出器
11日前
株式会社大真空
センサ
22日前
株式会社大真空
センサ
6日前
甲神電機株式会社
電流センサ
11日前
株式会社高橋型精
採尿具
12日前
ユニパルス株式会社
ロードセル
5日前
株式会社トプコン
測量装置
11日前
ダイトロン株式会社
外観検査装置
18日前
株式会社トプコン
測量装置
25日前
アズビル株式会社
漏液センサ
22日前
学校法人東京電機大学
干渉計
19日前
株式会社国際電気
治具セット
25日前
アズビル株式会社
熱式流量計
12日前
株式会社諸岡
自動運転作業機
22日前
豊田合成株式会社
表示装置
11日前
株式会社ミトミ技研
圧力測定装置
18日前
TDK株式会社
磁気センサ
18日前
日本特殊陶業株式会社
ガスセンサ
12日前
シャープ株式会社
収納装置
12日前
株式会社JVCケンウッド
撮像装置
19日前
理研計器株式会社
ガス検知器
22日前
個人
電気計器用結線器
22日前
エスペック株式会社
試験装置
19日前
トヨタ自動車株式会社
画像検査装置
6日前
株式会社ジェイテクト
荷重付与装置
25日前
アズビル株式会社
真空計測システム
22日前
株式会社東京久栄
水中移動体用採水器
12日前
アズビル株式会社
ヒータ温度推定装置
19日前
株式会社ミツトヨ
目盛板姿勢検査方法
25日前
エスペック株式会社
二次電池用プローブ
25日前
ローム株式会社
MEMSデバイス
12日前
株式会社エー・アンド・デイ
金属検出機
22日前
横河電機株式会社
光源装置
12日前
続きを見る
他の特許を見る




 特許ウォッチ
特許ウォッチ