TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
公開番号
2024105740
公報種別
公開特許公報(A)
公開日
2024-08-07
出願番号
2021082633
出願日
2021-05-14
発明の名称
ゲノム改変微生物株
出願人
国立大学法人 東京大学
,
独立行政法人酒類総合研究所
代理人
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
,
個人
主分類
C12N
1/19 20060101AFI20240731BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】糖をアルコールに代謝する微生物に対して、直列育種工程により2以上の遺伝子に変異を導入する工程を含む、遺伝子改変微生物を製造する方法を提供する。
【解決手段】単糖をアルコール類へ代謝する微生物を準備する工程と、前記微生物ゲノム上の2以上の遺伝子に対して、直列育種により変異を導入する工程とを含む、遺伝子改変微生物を製造する方法であって、前記2以上の遺伝子が、
a)高泡形成性遺伝子;
b)アルギナーゼ遺伝子;
c)メチオニンサルベージ経路の遺伝子;および
d)脂肪酸合成酵素のαサブユニットをコードする遺伝子、
から選択されてよく、前記遺伝子改変微生物が清酒酵母であってよい、方法である。
【選択図】図1
特許請求の範囲
【請求項1】
単糖をアルコール類へ代謝する微生物を準備する工程と、
前記微生物ゲノム上の2以上の遺伝子に対して、直列育種により変異を導入する工程と
を含む、遺伝子改変微生物を製造する方法。
続きを表示(約 870 文字)
【請求項2】
前記2以上の遺伝子が、
a)高泡形成性遺伝子;
b)アルギナーゼ遺伝子;
c)メチオニンサルベージ経路の遺伝子;および
d)脂肪酸合成酵素のαサブユニットをコードする遺伝子、
から選択される、請求項1に記載の伝子改変微生物を製造する方法。
【請求項3】
前記遺伝子改変微生物が清酒酵母である、請求項1または2に記載の方法。
【請求項4】
a)高泡形成性遺伝子;
b)アルギナーゼ遺伝子;
c)メチオニンサルベージ経路の遺伝子;または
d)脂肪酸合成酵素のαサブユニットをコードする遺伝子、
から選択される2以上の遺伝子に変異を有する、
糖をアルコールへ代謝する微生物。
【請求項5】
a)高泡形成性遺伝子がAWA1遺伝子またはその相同体であり、
b)アルギナーゼ遺伝子がCAR1遺伝子又はその相同体であり、
c)メチオニンサルベージ経路の遺伝子がMDE1遺伝子若しくはMRI1遺伝子又はこれらの相同体であり、
d)脂肪酸合成酵素のαサブユニットをコードする遺伝子がFAS2遺伝子又はその相同体である、
請求項4に記載の微生物。
【請求項6】
前記微生物が多倍体真核生物であり、前記変異がホモ接合性変異である、請求項4~5のいずれか一項に記載の微生物。
【請求項7】
前記微生物が清酒酵母である請求項4~6のいずれか一項に記載の微生物。
【請求項8】
前記微生物が清酒酵母協会6号、協会7号、協会9号、協会10号のいずれかである請求項7に記載の微生物。
【請求項9】
前記糖が、グルコースである請求項4~8のいずれか一項に記載の微生物。
【請求項10】
前記アルコールがエチルアルコールである、請求項4~9のいずれか一項に記載の微生物。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
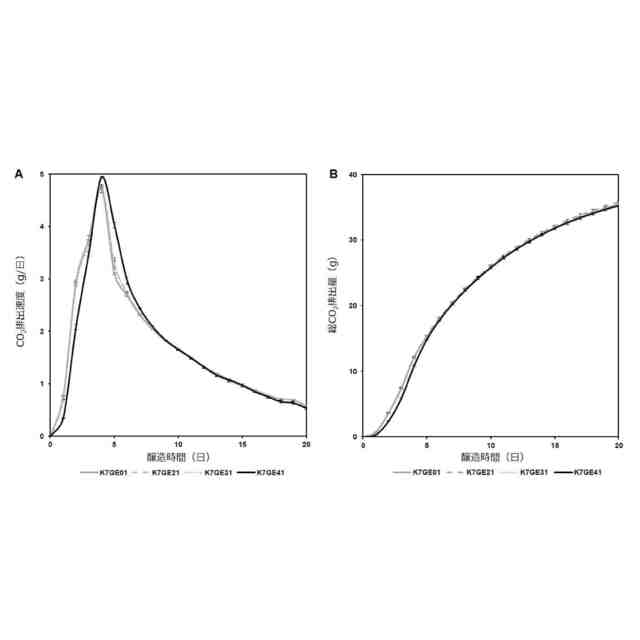
本開示は、直列育種工程により2以上の遺伝子に変異を導入した、糖をアルコールに代謝する遺伝子改変微生物株およびその使用、並びに糖をアルコールに代謝する微生物に対して、直列育種工程により2以上の遺伝子に変異を導入する工程を含む、遺伝子改変微生物を製造する方法に関する。
続きを表示(約 2,700 文字)
【背景技術】
【0002】
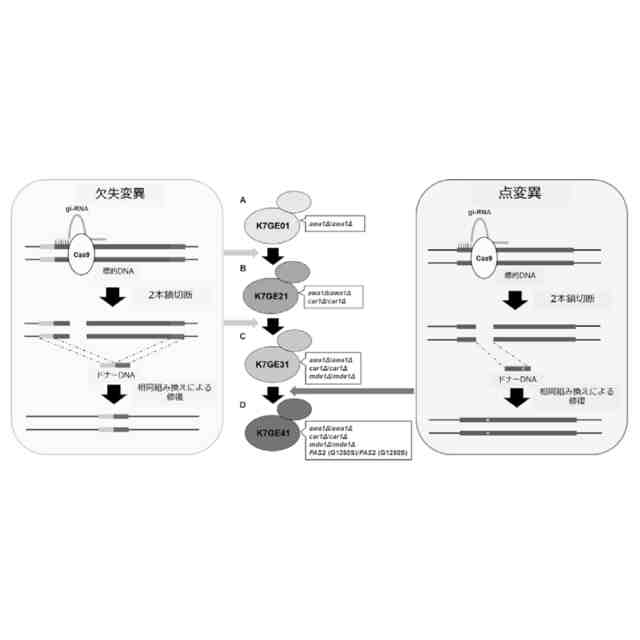
グルコースなどの糖をエチルアルコールなどのアルコールに代謝する微生物は醸造による酒類の製造等に広範に利用されてきた。これらの微生物は遺伝的背景の相違によって、製造中間物質及び最終製品の性質に様々な相違をもたらすことが知られている。これら製造中間物質の性質は製造工程の効率等に影響を与え、また、最終製品の性質は、例えば、製造される酒類の味、香り等に影響を与える。清酒を製造するのに使用される清酒酵母株では、もろみ中で分厚い泡の層(高泡)を形成しない酵母株が知られており、高い醸造効率をもたらす当該酵母株として利用されている。このような微生物の遺伝的背景と表現型の関係が研究された結果、醸造特性に重要ないくつかの遺伝子が明らかになってきた。これらの情報に基づいて遺伝子工学により、優れた特性を持つパン酵母株の構築が可能になってきた。例えば、代表的な清酒酵母きょうかい酵母(登録商標)7号(K7)に自然突然変異が導入された酵母株を交配、スクリーニングすることにより、高泡形成遺伝子AWA1が一部欠損した泡なし酵母K701などが作製されている(非特許文献1)。同様に、AWA1遺伝子が欠損した清酒酵母として、きょうかい酵母(登録商標)6号(K6)にAWA1欠損を導入した酵母K601、きょうかい酵母(登録商標)10号(K10)にAWA1欠損を導入した酵母K1001などが育種されている(並列育種)。
【0003】
しかしながら、このような交配とスクリーニングによる新規微生物育種株の取得は、意図された遺伝子変異以外の変異(オフターゲット変異)の蓄積を伴い得る。例えば、上記酵母K701については、ゲノム配列比較の結果、意図されたAWA1の一部欠損の他に、意図されない変異(オフターゲット変異)が蓄積していることが明らかになった(非特許文献2)。具体的には、K701はK7に対して、5個の遺伝子において73個のヘテロ接合性またはホモ接合性の非同義変異を有していた。育種株におけるオフターゲット変異の存在は親株の優良な醸造特性の損失につながることが懸念され、意図された変異以外の変異は最小限に抑制されることが好ましい。
【0004】
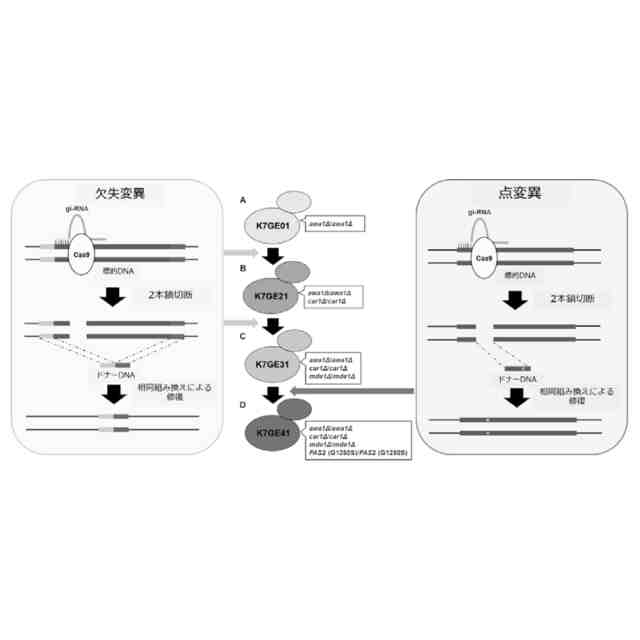
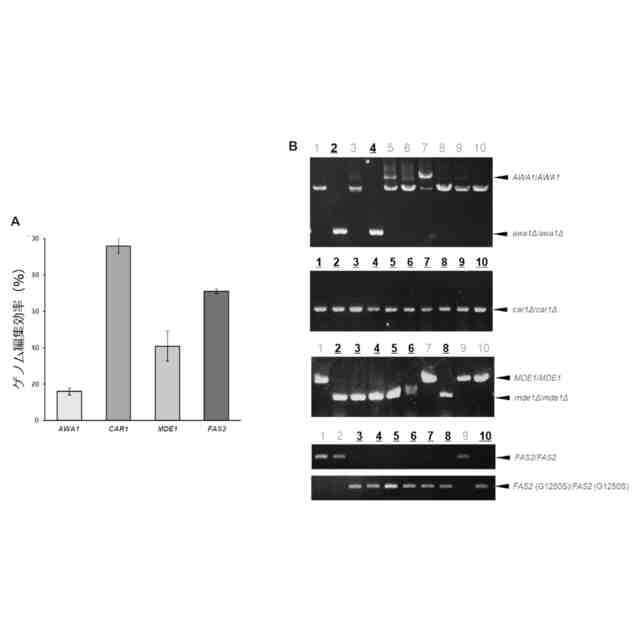
発明者らはオフターゲット変異を持たない理想的な清酒酵母育種の作製を目的として、CRISPR-Cas9システムを用いた清酒酵母の遺伝子破壊株の作製を行ってきた。その結果、世界にさきがけてK7から泡なし表現型を持つawa1Δ/awa1Δ株、K7GE01を単離した(非特許文献2、3)。
【0005】
一方、香りや有害物質の産生に関係する様々な遺伝子およびその変異が同定されており、これら複数の遺伝子に変異を有する微生物育種を製造することができれば、優れた特質を兼ね備えた新規微生物育種が取得されることが期待される。
【0006】
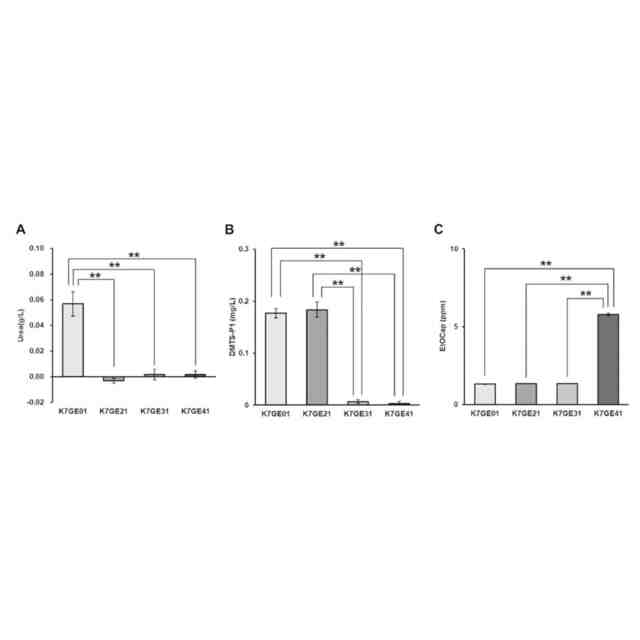
例えば、清酒醸造酵母が生成するジメチルトリスルフィド(DMTS)は、清酒を貯蔵した際に生じる好ましくない劣化臭「老香(ひねか)」の原因物質であることが知られている(非特許文献4)。清酒醸造酵母においてはDMTSの生成に酵母代謝系におけるメチオニンサルベージ経路が関与することが示唆されており、当該経路に関係するMDE1遺伝子またはMRI1遺伝子を欠失させることにより、DMTSの前駆体であるDMTS-P1の生成が著明に減少することが知られている(非特許文献5)。
【0007】
また、例えば清酒において好まれる果実香様吟醸香の主体は、清酒醸造酵母が生成するカプロン酸エチルと酢酸イソアミルであることが知られている(非特許文献4)。清酒醸造酵母においては、脂肪酸合成酵素のαサブユニットをコードするFAS2遺伝子おいて1250番目のグリシンをセリンに置換した変異(Gly1250Ser変異)を有する変異株は、カプロン酸エチルの産生が増強され、強い吟醸香を有することが知られている(非特許文献6)。
【0008】
また、例えば清酒醸造酵母が生成する尿素からは、清酒の保存時に発癌性物質であるカルバミン酸エチルが生じるため、清酒醸造酵母による尿素生成を抑制することが好ましい。清酒酵母ではアルギナーゼ遺伝子(CAR1)を欠失させることで、尿素生成が抑制されることが知られている(非特許文献7)。
【0009】
しかしながら、遺伝子工学的手法を利用したとしても、これら複数の遺伝子のそれぞれに変異を有する微生物株を作成し、これらを交配とスクリーニング通じて複数の変異を同時に有する新規微生物育種株の取得する方法(直列育種)は、異なる親株に単一の変異を導入する並行育種に比して多大な時間を要することとなる。また、清酒酵母のように多倍体の微生物の場合には、劣性突然変異を導入する場合にホモ接合対立遺伝子を準備する必要がある。さらに、清酒酵母のように遺伝子組み換えの生じた微生物株を選択するためのマーカーが存在しない微生物では、直列育種によって、複数の遺伝子に意図された変異を同時に有する微生物株を製造することは極めて困難であった。
【先行技術文献】
【非特許文献】
【0010】
K.Ouchi,and H.Akiyama,Agric.Biol.Chem.,1971,Vol.35,pp.1024-1032
Ohnuki S et Al.,Biosci Biotechnol Biochem,2019,Vol.83,no.8,pp.1-11
山田駿一 修士論文2018
科学と教育、2015、Vol.63,No.10,pp.506-507
Makimoto J et al.,Journal of Bioscience and Bioengineering,2020,Vol.130,No.6,pp.610-615
Ichikawa,E.et al.,Agric. Biol. Chem.,1991,Vol.55,no.8,pp.2153-2154
Kitamoto K,et al.,Appl Environ Microbiol,1991,Vol.57,no.1,pp.301-306
【発明の概要】
【発明が解決しようとする課題】
(【0011】以降は省略されています)
特許ウォッチbot のツイートを見る
この特許をJ-PlatPatで参照する
関連特許
国立大学法人 東京大学
プロペラ
2日前
国立大学法人 東京大学
情報処理装置
2か月前
国立大学法人 東京大学
腸管壁保持装置
1か月前
国立大学法人 東京大学
ロボットアーム
1か月前
国立大学法人東京農工大学
タンクモジュール
25日前
国立大学法人 東京大学
双方向散乱顕微鏡
1か月前
国立大学法人 東京大学
てんかんの検出方法
1か月前
国立大学法人 東京大学
アンモニア製造方法
1か月前
国立大学法人 東京大学
化合物半導体チップ
1か月前
国立大学法人 東京大学
粘性の測定装置及び方法
18日前
国立大学法人 東京大学
アシルヒドラゾン誘導体
2か月前
国立大学法人 東京大学
制御装置、および制御方法
23日前
国立大学法人 東京大学
消化管幹細胞活性化用組成物
2日前
国立大学法人 東京大学
海産魚スクーチカ症予防治療剤
1か月前
国立大学法人 東京大学
試料測定装置及び試料測定方法
1か月前
国立大学法人 東京大学
炭酸カルシウム結晶の製造方法
2日前
ダイキン工業株式会社
金属錯体
1か月前
国立大学法人 東京大学
血糖制御能力の推定方法及び装置
25日前
国立大学法人 東京大学
切片作成装置および切片作成方法
1か月前
国立大学法人 東京大学
搬送トレイおよびワーク作業装置
1か月前
国立大学法人 東京大学
スピーカー及びオーディオシステム
11日前
ダイキン工業株式会社
薄膜の製造方法
1か月前
ダイキン工業株式会社
薄膜の製造方法
1か月前
住友ゴム工業株式会社
錯体の製造方法
1か月前
東京瓦斯株式会社
液体肥料の製造方法
1か月前
国立大学法人 東京大学
情報処理装置、制御装置、及びプログラム
2か月前
三洋化成工業株式会社
熱化学電池用電解液
2か月前
国立大学法人 東京大学
SiCウェハのダイシング方法及びその装置
2か月前
国立大学法人 東京大学
東大式メイクアップ理論による肌年齢推定方法
2日前
国立大学法人 東京大学
情報処理システム、プログラム及び情報処理方法
24日前
国立大学法人 東京大学
ジエン系高靭性エラストマーおよびその製造方法
1か月前
国立大学法人 東京大学
伸縮性配線基板及び該伸縮性配線基板の製造方法
1か月前
国立大学法人 東京大学
情報処理システム、プログラム及び情報処理方法
24日前
ダイキン工業株式会社
検知方法および検知装置
1か月前
国立大学法人 東京大学
情報処理装置、情報処理システム、及びプログラム
1か月前
国立大学法人 東京大学
プログラム、情報処理システム、及び情報処理方法
2か月前
続きを見る
他の特許を見る

 特許ウォッチ
特許ウォッチ