TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025168349
公報種別
公開特許公報(A)
公開日
2025-11-07
出願番号
2025124128,2022525402
出願日
2025-07-24,2020-11-16
発明の名称
TMEM219抗体及びその治療的使用
出願人
エンテラ・エッセ・エッレ・エッレ
代理人
個人
,
個人
主分類
C07K
16/28 20060101AFI20251030BHJP(有機化学)
要約
【課題】高い親和性でヒトecto-TMEM(TMEMの細胞外ドメイン)に特異的に結合し、結合の際にTMEM219経路を活性化することなく、その同族の受容体であるTMEM219へのIGFBP3の結合を低減又は排除することができる抗体を提供すること。
【解決手段】請求項1に記載のヒトTMEM219受容体に結合し、前記TMEM219受容体へのインスリン様成長因子3(IGFBP3)の結合を阻害又は低減する、単離された抗体又はその抗原結合性断片により、上記課題を解決する。
【選択図】なし
特許請求の範囲
【請求項1】
ヒトTMEM219受容体に結合し、前記TMEM219受容体へのインスリン様成長因子3(IGFBP3)の結合を阻害又は低減する、単離された抗体又はその抗原結合性断片であって、
a.以下を含む重鎖可変ドメイン(VH):
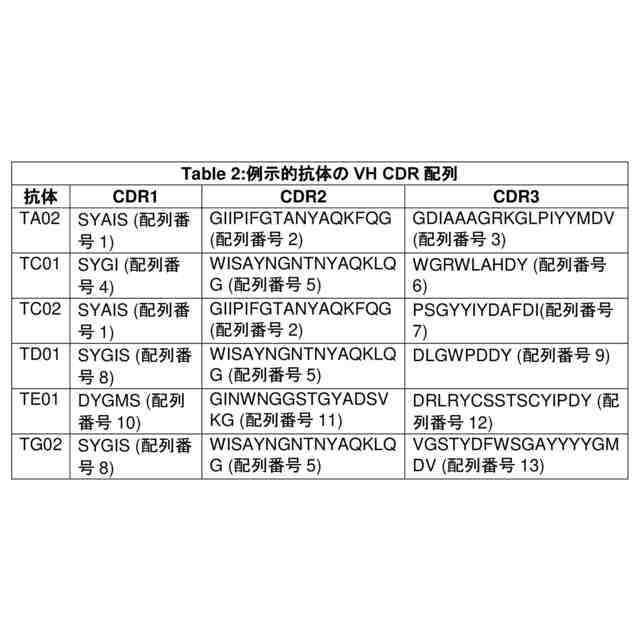
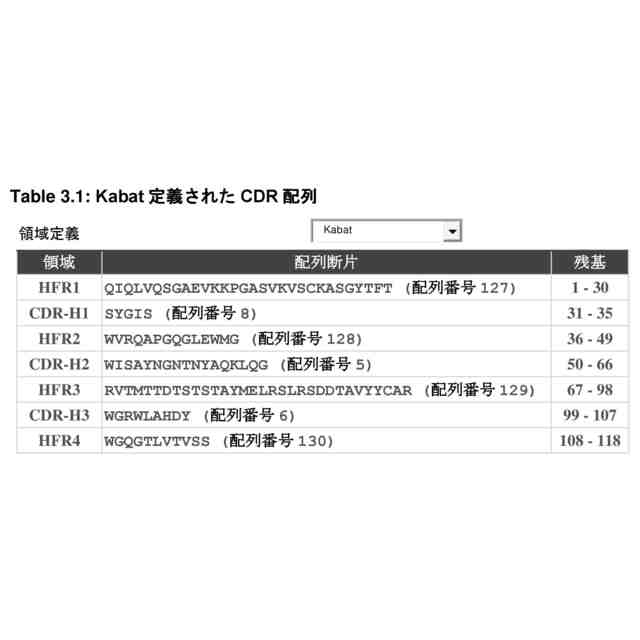
i.配列番号1、8、10、56、59、62、65、及び68からなる群から選択されるアミノ酸配列のCDR1配列;
ii.配列番号5、2、11、57、60、63、66、及び69からなる群から選択されるアミノ酸配列のCDR2配列;及び
iii.配列番号3、7、9、12、13、58、61、64、67、及び70からなる群から選択されるアミノ酸配列のCDR3配列;並びに
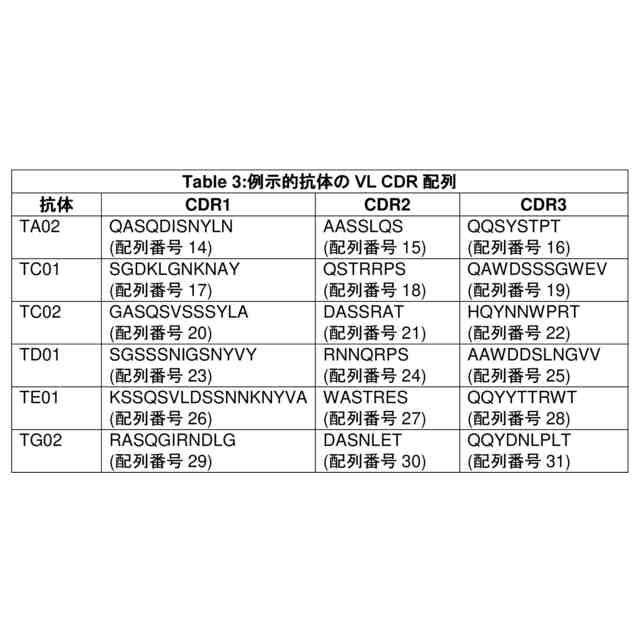
b.以下を含む軽鎖可変ドメイン(VL):
i.配列番号14、20、23、26、29、71、77、80、82、及び85からなる群から選択されるアミノ酸配列のCDR1配列;
ii.配列番号15、21、24、27、30、72、78、83、及び86からなる群から選択されるアミノ酸配列のCDR2配列;及び
iii.配列番号16、22、25、28、31、73、74、75、76、79、81、84、及び87からなる群から選択されるアミノ酸配列のCDR3配列
を含む、単離された抗体又はその抗原結合性断片。
続きを表示(約 2,200 文字)
【請求項2】
- 配列番号1、配列番号2、配列番号7、配列番号20、配列番号21、及び配列番号22;
- 配列番号1、配列番号2、配列番号3、配列番号14、配列番号15、及び配列番号16;
- 配列番号8、配列番号5、配列番号9、配列番号23、配列番号24、及び配列番号25;
- 配列番号10、配列番号11、配列番号12、配列番号26、配列番号27、及び配列番号28;
- 配列番号8、配列番号5、配列番号13、配列番号29、配列番号30、及び配列番号31;
- 配列番号56、配列番号57、配列番号58、配列番号71、配列番号72、及び配列番号73;
- 配列番号56、配列番号57、配列番号58、配列番号71、配列番号72、及び配列番号74;
- 配列番号56、配列番号57、配列番号58、配列番号71、配列番号72、及び配列番号75;
- 配列番号59、配列番号60、配列番号61、配列番号71、配列番号72、及び配列番号76;
- 配列番号56、配列番号57、配列番号58、配列番号77、配列番号78、及び配列番号79;
- 配列番号62、配列番号63、配列番号64、配列番号80、配列番号78、及び配列番号81;
- 配列番号65、配列番号66、配列番号67、配列番号82、配列番号83、及び配列番号84;又は
- 配列番号68、配列番号69、配列番号70、配列番号85、配列番号86、及び配列番号87;
を含む、請求項1に記載の単離された抗体又はその抗原結合性断片。
【請求項3】
IGFBP3の結合によって誘導されるTMEM219受容体の活性化を阻害、低減、又は中和する、及び/又は、
ヒトTMEM219受容体に結合する際にTMEM219経路を活性化しない、
請求項1又は2に記載の単離された抗体又はその抗原結合性断片。
【請求項4】
糖尿病患者においてβ細胞を保存すること、及び/又は膵島破壊を防ぐこと、及び/又はインビボモデルにおいて血糖値を制御することにおいて有効である、並びに/又は、
インビボモデルにおいて急性結腸炎を低減させることにおいて有効である、
請求項1から3のいずれか一項に記載の単離された抗体又はその抗原結合性断片。
【請求項5】
a-IGFBP3で処置された健康な対象のミニ腸成長の増加;
b-IBD患者のミニ腸成長の増加;
c-糖尿病性腸疾患血清で処置された健康な対象のミニ腸成長の増加;
d-IGFBP3で処置された健康な対象のミニ腸におけるEphB2及び/又はLGR5の発現の増加;
e-IGFBP3で処置された健康な対象のミニ腸におけるカスパーゼ8発現の減少;
から選択される少なくとも1つの活性を有する、請求項1から4のいずれか一項に記載の単離された抗体又はその抗原結合性断片。
【請求項6】
f-IGFBP3で処置されたβ細胞におけるβ細胞喪失の減少;
g-IGFBP3で処置されたβ細胞におけるインスリンの発現の増加;
j-IGFBP3で処置されたβ細胞におけるβ細胞のアポトーシスの減少;
から選択される少なくとも1つの活性を有する、請求項1から4のいずれか一項に記載の単離された抗体又はその抗原結合性断片。
【請求項7】
h-DSSで誘導された腸内細胞アポトーシスを阻害又は減少させる;
i-DSSで処置された結腸におけるPCNAの発現を回復させる;
から選択される少なくとも1つの活性を有する、請求項1から4のいずれか一項に記載の単離された抗体又はその抗原結合性断片。
【請求項8】
k-糖尿病の動物モデルにおける膵島炎スコアの減少;
l-糖尿病の動物モデルにおける糖尿病の発症の減少;
m-糖尿病の動物モデルにおけるβ細胞傷害を保護する;
n-糖尿病の動物モデルにおけるβ細胞喪失を防ぐ
から選択される少なくとも1つの活性を有する、請求項1から4のいずれか一項に記載の単離された抗体又はその抗原結合性断片。
【請求項9】
a)、b)、及びc)における増加が、少なくとも20%である;d)における増加及びe)における減少が、少なくとも30%である;又はf)における減少及びg)における増加が、少なくとも10%である、請求項5又は6に記載の単離された抗体又はその抗原結合性断片。
【請求項10】
a.配列番号32、配列番号34~配列番号37、及び配列番号88~配列番号95からなる群から選択されるアミノ酸配列の重鎖可変ドメイン(VH)配列;及び
b. 配列番号38、配列番号40~配列番号43、及び配列番号96~配列番号170からなる群から選択されるアミノ酸配列の軽鎖可変ドメイン(VL)配列
を含む、請求項1から9のいずれか一項に記載の単離された抗体又はその抗原結合性断片。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、IGFBP3受容体、すなわちTMEM219に特異的に結合する抗体又はその抗原結合性断片、それらの生産方法、前記抗体を含む医薬組成物、及びその使用に関する。
続きを表示(約 3,200 文字)
【背景技術】
【0002】
IGFBP3/TMEM219軸
インスリン様成長因子結合タンパク質は、インスリン様成長因子(IGF)のバイオアベイラビリティを調節する7つの結合タンパク質のファミリーである。それらの中でも、IGFBP3は最も豊富であり、ほとんど全ての組織に存在し、IGFに対してより高い親和性を有し;実際に、IGFのおよそ80~90%は、酸不安定性サブユニット(ALS)との三元複合体におけるIGFBP3に結合する(1)。
【0003】
IGFのアベイラビリティを調節するその能力に加えて、IGFBP3は、IGF非依存性機能も有することが分かっている(2)。実際に、それは、細胞表面タンパク質、必須のシグナル伝達能力を有する細胞表面受容体、細胞内タンパク質及び核タンパク質(転写調節因子)と会合することができ、結果として、細胞増殖に影響を及ぼし、アポトーシスを直接誘導する(2)。死受容体の中でも、シングルスパン膜タンパク質であるTMEM219は、IGFBP-3への高い結合性が示された(3)。TMEM219へのIGFBP3の結合はがん細胞(すなわち、前立腺がん細胞及び乳がん細胞)を含む様々な細胞においてカスパーゼ8媒介性アポトーシスを誘導するが(3)、幹細胞(すなわち、結腸幹細胞)においても誘導する(4)。様々な戦略によってIGFBP3/TMEM219軸を遮断又は増強することは、それぞれ細胞死を防止又は増加させることが分かっている。発明者らの知る限りにおいて、IGFBP3/TMEM219の結合を防ぐことができ、TMEM219へのIGFBP3の結合の標的組織/細胞に対するIGF-I非依存性のカスパーゼ8媒介性有害効果を止めることができるような、市販のTMEM219又はIGFBP3に対するモノクローナル抗体は存在しない。
【0004】
糖尿病におけるIGFBP3/TMEM219軸
1型(T1D)及び2型糖尿病(T2D)は両方とも、結果としてインスリンの分泌の低減、血糖値の制御不全、及び多糖症を引き起こすβ細胞の喪失によって特徴付けられる(5、6)。異なる発症メカニズムにもかかわらず、T1Dにおける自己免疫反応又はT2Dにおけるインスリン耐性/炎症は、両方とも、β細胞集団の漸減を生じる。実際に、発生する自己免疫活性化は、T1Dにおけるβ細胞喪失を完全に説明するのに十分なようには思われないということが明らかになりつつある(5)。その上、免疫療法ではT1Dを治療することができない(7)ことは、(i)自己免疫は、T1D病因に関与する唯一の要因ではないかもしれないこと、及び、(ii)T1Dに対する効果的な処置を確立するためには、β細胞喪失等の疾患の異なるメカニズムを標的にする代替戦略が必要であることを浮き彫りにした。点在するβ細胞が長期のT1Dを有する個体において検出されるという観察(8)は、β細胞の代謝回転を維持するために新たなβ細胞が生じなければならないか(5、9)、又は破壊されたβ細胞は、「異なり」得るか、又は死ぬ傾向にあり得るか(10)のどちらかであることを裏付けている。このことは、表面β細胞受容体の上方/下方制御された発現が、それらを免疫システムに認識させるために、重要な役割を有し得ること、より重要なことに、他の非免疫学的決定因子が、β細胞の運命及び機能を調節し得ることを示唆し得る。したがって、T1Dにおける非免疫学的β細胞破壊及びT2Dにおけるβ細胞の進行性喪失を防ぐことは、β細胞の発生と破壊の間のバランスを、適切なβ細胞集団の回復へと傾け得、結果として、疾患の最初の段階を停止又は遅延させることができる新規の治療アプローチのための道を整え得る。IGFBP3受容体であるTMEM219は、β細胞系及びヒト/マウス膵島において発現されること、及びそのライゲーションは、β細胞に対して毒性であることが分かっている。興味深いことに、ヒトIGFBP3に対するマウス遺伝子移植は、多糖症を発症し、膵島質量の減少を示し、インスリン-グルコース刺激に対する反応の低減を示すが(11)、その一方で、IGFBP3のためにノックダウンされたものは、糖代謝制御に関するいかなる変更も示さなかった(12)ことも観察されている。
【0005】
ヒトにおいて、最近、Drogan及び同僚は、IGFBP3の高い血中濃度が、T2Dの発症に関連することを公開した(13)。その上、Diabimmune研究グループによる最近の研究は、IGFBP3レベルが、T1Dに対するリスクにある子供での血清変換に対する自己抗体の陽性及び機会と相関し、したがって、β細胞自己免疫の早期発達におけるIGFBP3の循環の役割を示唆していることを実証した(14)。
【0006】
IGFBP3受容体であるTMEM219は、死受容体として既に記載されているが、その活性化は、標的細胞内においてカスパーゼ8媒介性アポトーシスを引き起こし、結果として、それらの喪失を生じる(4)。
【0007】
炎症性腸疾患におけるIGFBP3/TMEM219軸
腸幹細胞(ISC)は、小腸及び大腸の陰窩の底部に存在し、陰窩再生及び代謝回転を制御する。特に、ISCは、陰窩に沿って分化することにより、杯細胞、腸細胞、腸内分泌細胞を生じることができる(4)。
【0008】
炎症性腸疾患(IBD)は、2つの臨床エンティティであるクローン病(CD)及び潰瘍性大腸炎(UC)を包含する免疫媒介性の慢性状態であり、ヨーロッパの約250万人及び米国の100万人に影響を及ぼす(15)。IBDの病因は、まだ調査中であるが、最近の証拠は、回腸CDにおけるパーネト細胞への、並びにUCにおける杯細胞へのISCの分化の減少が、当該疾患の発症において重要な役割を果たし得ることを示唆している。特に、粘膜での局所シグナル伝達及び炎症性経路は両方とも、外部刺激に反応して、ISCの数及び機能を保ち、結果として、腸の恒常性を維持する(16)。実際に、近年、Yancuらは、CDにおけるIGFBP-3の役割を支持する結果を公開した。実際に、彼らは、デキストラン-ナトリウム-サルフェート(DSS)大腸炎マウスモデルにおいて、IGFBP3のノックアウトが、炎症を調節する役割を有することを実証した(17)。
【0009】
本発明者らは、最近、インスリン様成長因子結合タンパク質3(IGFBP3)受容体、すなわち、TMEM219受容体が、ISCにおいて発現されること、並びに循環ホルモンIGFBP3との相互作用が、糖尿病及び糖尿病性腸疾患での腸障害のモデルにおいて、ISCの運命及び機能を制御することを見出した(4)。糖尿病性腸疾患及びIBDは、腸幹細胞(ISC)恒常性における変更及び粘膜モルホロジーの変更として、共通の特徴を共有するため、これらの結果は、依然として未知であるIBD病因における重要な洞察を加え得、おそらく、IBD処置のための新規の治療アプローチの導入につながるであろう。
【0010】
IBDに対して現在利用可能な療法は、抗炎症剤の使用及び免疫療法的戦略に基づいており、それらは、いくつかの有害作用及び長期での有効性に対する疑わしさによって悪化される。特にUCでは、疾患の進行した状態において、外科手術も首尾よく用いられている(15)。主にCDにおける疾患の再発も頻繁であり、結果として、異なる治療アプローチの必要性を浮き彫りにする。結果として、IBDの処置における新規の治療標的及び治療戦略の特定は、臨床的に非常に重要であり、健康な共同体にとって必要である。
(【0011】以降は省略されています)
この特許をJ-PlatPat(特許庁公式サイト)で参照する
関連特許
エンテラ・エッセ・エッレ・エッレ
TMEM219抗体及びその治療的使用
3日前
クミアイ化学工業株式会社
多形
11日前
東ソー株式会社
タンパク質の発現方法
1か月前
東ソー株式会社
炭素-窒素結合形成方法
4か月前
株式会社トクヤマ
四塩化炭素の製造方法
4か月前
株式会社トクヤマ
シロキサン類の回収方法
4か月前
株式会社半導体エネルギー研究所
有機化合物
2か月前
株式会社トクヤマ
ビオチン誘導体の製造方法
3か月前
株式会社半導体エネルギー研究所
有機化合物
11日前
栗田工業株式会社
ギ酸の回収方法
1か月前
株式会社トクヤマ
ホルムアミド化合物の製造方法
2か月前
株式会社トクヤマ
チオラクトン誘導体の製造方法
1か月前
株式会社コスモス
液状炭化水素の増産方法
3か月前
株式会社コスモス
液状炭化水素の増産方法
3か月前
東ソー株式会社
イソシアネート化合物の製造方法
2か月前
株式会社トクヤマ
ベンザゼピン化合物の製造方法
3か月前
株式会社トクヤマ
チオファニウム塩誘導体の製造方法
13日前
信越化学工業株式会社
新規化合物
4か月前
artience株式会社
四塩基酸無水物の製造方法
4か月前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
3か月前
国立大学法人東京農工大学
深共晶溶媒
2か月前
三菱ケミカル株式会社
アルコールの製造方法
2か月前
大阪瓦斯株式会社
メタン製造システム
1か月前
本田技研工業株式会社
CO2変換方法
1か月前
株式会社トクヤマ
アシル化ベンゼン誘導体の酸塩の製造方法
2か月前
JNC株式会社
有機ケイ素化合物およびこれを用いた重合体
2か月前
JNC株式会社
有機ケイ素化合物およびこれを用いた重合体
3か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
1か月前
キヤノン株式会社
有機化合物及び有機発光素子
1か月前
株式会社半導体エネルギー研究所
有機金属錯体、発光デバイス
2か月前
キヤノン株式会社
有機金属錯体及び有機発光素子
1か月前
個人
IL-17産生誘導能を有する化合物及びその用途
1か月前
国立大学法人京都大学
抗がん剤
2か月前
デンカ株式会社
有機酸又はその塩の製造方法
2か月前
国立大学法人岩手大学
反応性付与化合物
2か月前
株式会社成和化成
アスコルビン酸誘導体又はその塩及びその化粧料
24日前
続きを見る
他の特許を見る



 特許ウォッチ
特許ウォッチ