TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025168346
公報種別
公開特許公報(A)
公開日
2025-11-07
出願番号
2025123833,2022518227
出願日
2025-07-24,2020-09-18
発明の名称
DNA塩基編集のための方法及び組成物
出願人
シンジェンタ クロップ プロテクション アクチェンゲゼルシャフト
代理人
個人
,
個人
,
個人
,
個人
,
個人
,
個人
主分類
C07K
19/00 20060101AFI20251030BHJP(有機化学)
要約
【課題】細胞のゲノム中の標的部位を改変するための方法及び組成物を提供すること。
【解決手段】改善されたリンカー配列で接続されている1個又は複数個のDNA結合ドメイン及び1個又は複数個の異種ドメイン、例えばDNA改変ドメインを含む、融合タンパク質を提供する。改善されたリンカー配列により接続されている1個又は複数個のDNA結合ドメイン及び1個又は複数個の異種ドメインを含む融合タンパク質をコードする、コドン最適化ポリヌクレオチドを提供する。
【選択図】なし
特許請求の範囲
【請求項1】
N末端からC末端の方向に、異種ドメイン、第1のリンカー配列、及びV型CRISPR-Cas酵素を含む融合タンパク質であって、前記第1のリンカー配列が、反復GGGGS配列を含む、融合タンパク質。
続きを表示(約 750 文字)
【請求項2】
前記異種ドメインが、デアミナーゼ、ポリメラーゼ、ヌクレアーゼ、リラクサーゼ、アルキルトランスフェラーゼ、メチルトランスフェラーゼ、アデノシンデアミナーゼ、シチジンデアミナーゼ、オキシダーゼ、チミンアルキルトランスフェラーゼ、アデニンオキシダーゼ、アデノシンメチルトランスフェラーゼ、グリコシラーゼ、又は核局在化シグナルである、請求項1に記載の融合タンパク質。
【請求項3】
前記異種ドメインが、デアミナーゼドメインである、請求項2に記載の融合タンパク質。
【請求項4】
前記デアミナーゼドメインが、シチジンデアミナーゼである、請求項3に記載の融合タンパク質。
【請求項5】
前記シチジンデアミナーゼドメインが、活性化誘導シチジンデアミナーゼ(「AID」)である、請求項4に記載の融合タンパク質。
【請求項6】
前記シチジンデアミナーゼドメインが、アポリポタンパク質B mRNA編集複合体(「APOBEC」)ドメインである、請求項4に記載の融合タンパク質。
【請求項7】
前記APOBECドメインが、APOBEC1ファミリのデアミナーゼである、請求項6に記載の融合タンパク質。
【請求項8】
前記APOBECドメインが、配列番号1と少なくとも70%同一の配列を含む、請求項7に記載の融合タンパク質。
【請求項9】
前記デアミナーゼドメインが、アデニンデアミナーゼである、請求項3に記載の融合タンパク質。
【請求項10】
前記アデニンデアミナーゼが、TadAドメインである、請求項9に記載の融合タンパク質。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、細胞のゲノム中における標的ヌクレオチド塩基編集のための方法及び組成物に関する。
続きを表示(約 3,300 文字)
【0002】
配列表の電子的提出に関する陳述
本明細書には、2020年9月18に作成されて37 C.F.R.§1.821に基づいて提出された、約702キロバイトの、「81945_ST25」という表題のASCIIテキスト形式の配列表が添付されており、且つ本明細書と共に提出されており、この配列表は、参照により本明細書に組み込まれる。
【背景技術】
【0003】
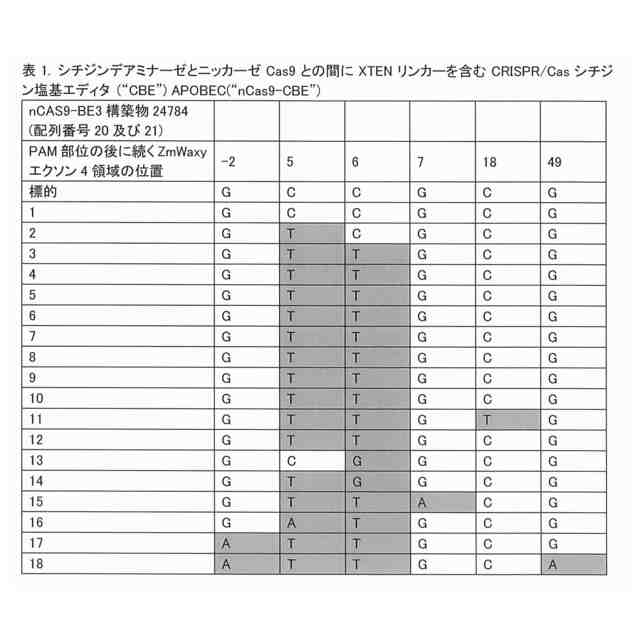
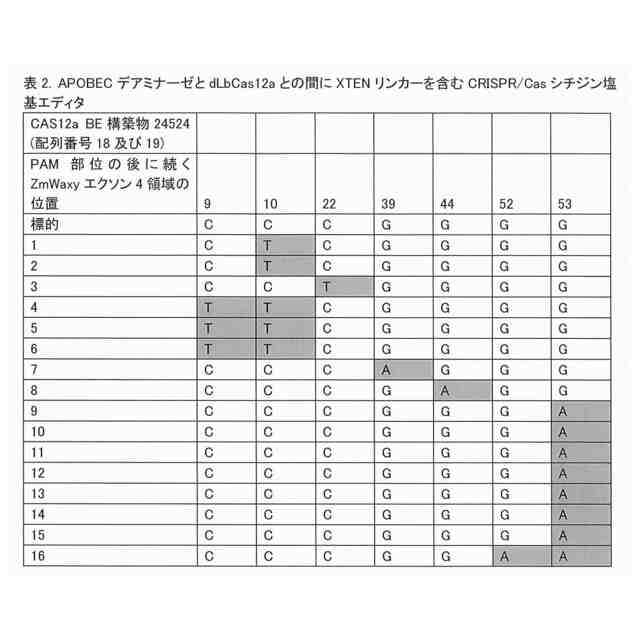
農業では、植物のゲノムを編集して有利な対立遺伝子を作り出す能力が非常に求められている。この能力により、収量が増加したり疾患が予防されたりする可能性がある。ゲノム編集は、植物では進展が遅れている新しい分野である。さらに、意図された変更以外のゲノムに対する変更には、望ましい変更の適用が制限されるという問題がある。CRISPR-CAS9は、DNAに二本鎖切断を行なうことにより機能する。この切断は、非相同末端結合又は相同性依存性修復により修復されることから、DNA塩基の挿入又は欠失が起こる場合がある。塩基編集と称される戦略により、切断されて挿入及び欠失が生じることなくDNAが変更される。1つのバージョンでは、シチジンデアミナーゼと称される酵素は、DNAを切断し得ないように改変されているCAS9酵素(Shimatani et al,2017.Nat.Biotechnol.35,441-443)又はCAS12a酵素(Li et al,2018.Nat.Biotechnol.36,324-327)により、特定の塩基を標的とする。シチジンデアミナーゼ及びヌクレアーゼ欠損CAS9又はCAS12aは、アミノ酸リンカーを介した接続により互いに融合している。リンカー接続の改善により、例えば、オフターゲット塩基変更が減少して切断の精度が改善され、これにより、この融合タンパク質の機能が改善され得る。
【発明の概要】
【課題を解決するための手段】
【0004】
この改善に関する要求を満たすために、本発明者らは、最適化されており且つ改善されているCas12a酵素及び構築物を提供する。具体的には、本発明者らは、異種ドメイン、第1のリンカー配列、及びV型CRISPR-Cas酵素を含む融合タンパク質を提供する。第1のリンカー配列は、反復GGGGS配列を含む。異種ドメインは、デアミナーゼ、ポリメラーゼ、ヌクレアーゼ、リラクサーゼ(relaxase)、アルキルトランスフェラーゼ、メチルトランスフェラーゼ、アデノシンデアミナーゼ、シチジンデアミナーゼ、オキシダーゼ、チミンアルキルトランスフェラーゼ、アデニンオキシダーゼ、アデノシンメチルトランスフェラーゼ、グリコシラーゼ、又は核局在化シグナルであり得る。塩基編集のために、異種ドメインは、デアミナーゼドメイン(例えば、シチジンデアミナーゼ又はアデニンデアミナーゼ)である。シチジンデアミナーゼドメインは、活性化誘導シチジンデアミナーゼ(「AID」)、又は例えばデアミナーゼのAPOBEC1ファミリ由来のアポリポタンパク質B mRNA編集複合体(「APOBEC」)ドメインであり得る。状況によっては、APOBECドメインは、配列番号1と少なくとも70%同一の配列を含む。アデニンデアミナーゼが必要な場合には、アデニンデアミナーゼは、配列番号92と少なくとも70%同一のアミノ酸配列を含むTadAドメインであり得る。
【0005】
V型CRISPR-Cas酵素がV-A型(「Cas12a」)酵素である場合には、Cas12aは、配列番号3、配列番号6、配列番号22、配列番号45、配列番号46、配列番号47、及び配列番号48で構成された群から選択される。Cas12aドメインは、触媒的に不活性であり得るが、標的DNAに依然として結合して異種ドメインが作動することを可能にする。Cas12aは不活性であり、この配列は、配列番号3、配列番号6、又は配列番号22である。
【0006】
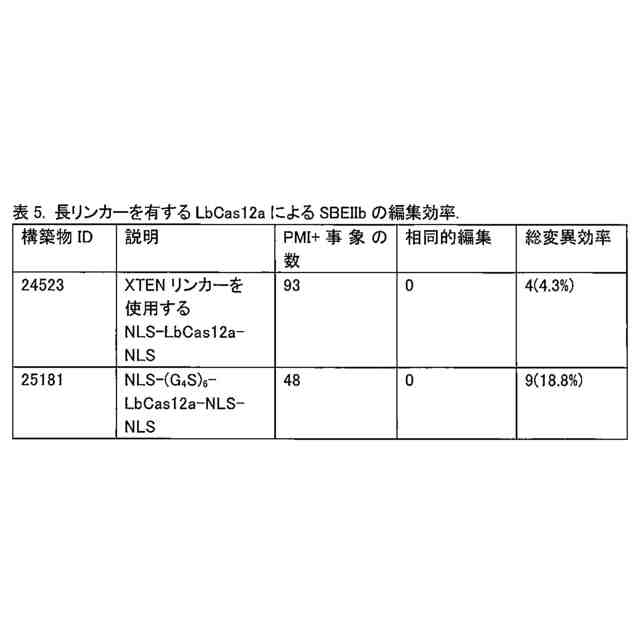
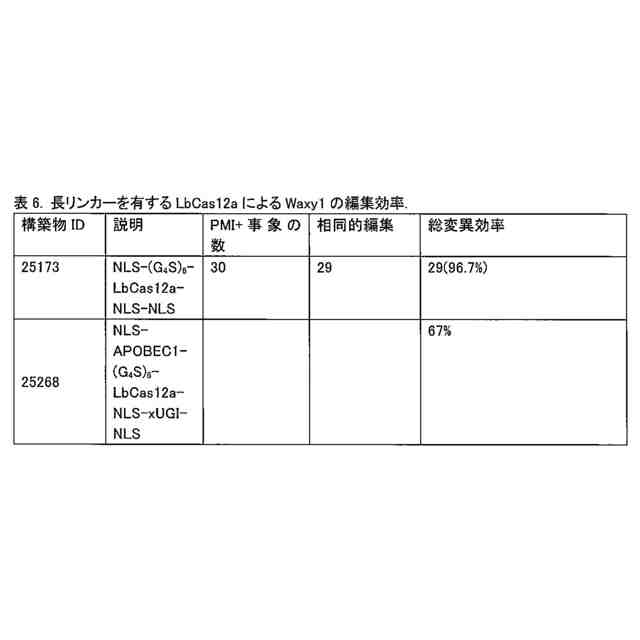
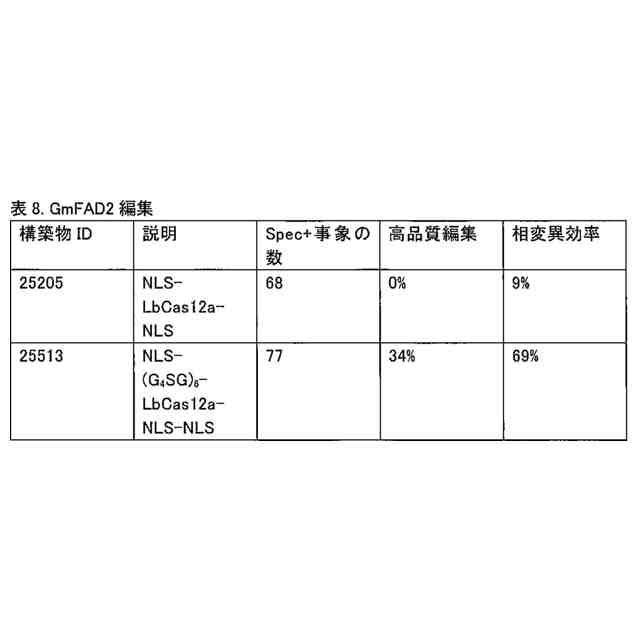
異種ドメインとCas12a酵素との間の第1のリンカー配列は、少なくとも3回反復されたGGGGSを含み得る。他の用途では、第1のリンカー配列は、少なくとも6回反復されたGGGGSを含み得る。
【0007】
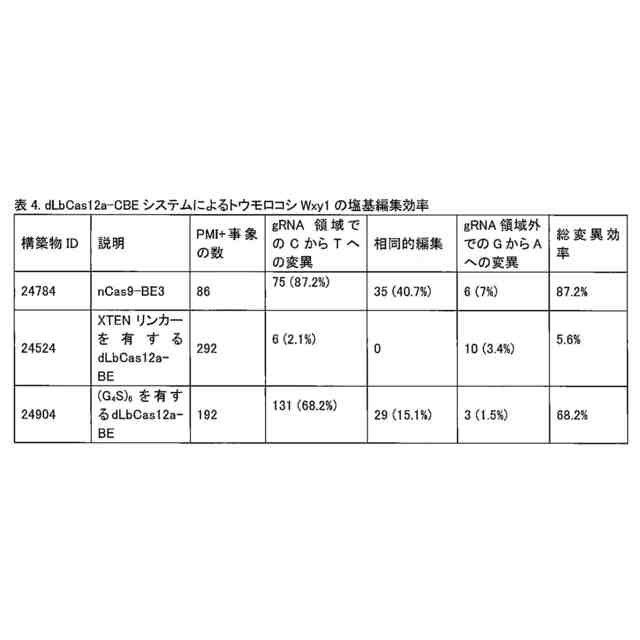
本融合タンパク質は、配列番号11、12、13、又は44を含み得、且つウラシルDNAグリコシラーゼインヒビター(「UGI」)ドメイン(配列番号8により表される)も含み得る。このUGIドメインは、配列SGGSを含む第2のリンカーによりCas12a酵素に連結され得る。この融合タンパク質は、配列番号17、配列番号24、配列番号35、配列番号39、配列番号43、配列番号50、配列番号52、配列番号54、配列番号56、配列番号81、配列番号83、配列番号85、配列番号87、又は配列番号89を含み得る。これらの融合タンパク質は、DNAと接触した場合には、反復GGGGS配列の第1のリンカー配列を欠く先行技術の融合タンパク質と比較して、高い頻度でオンターゲット編集を生じ、及び低い頻度でオフターゲット編集を生じる。
【0008】
本発明者らはまた、植物ゲノムDNAを編集する方法であって、植物ゲノムDNAと、(a)UGIドメインを任意に含む、上記態様の内の1つにより説明されている融合タンパク質、及び(b)工程(a)の融合タンパク質の標的を植物ゲノムDNAの標的DNA配列に定めるガイドRNA(「gRNA」)とを接触させることによる方法であり、編集された植物ゲノムDNAは、反復GGGGS配列以外の第1のリンカーを有する融合タンパク質により編集された植物ゲノムDNAと比較してオフターゲット編集が低い、方法も提供する。
【0009】
本発明者らはまた、低いオフターゲット編集で植物ゲノムDNAを編集する方法であって、植物ゲノムDNAと、(a)UGIドメインを任意に含む、上記態様の内の1つにより説明されている融合タンパク質、及び(b)工程(a)の融合タンパク質の標的を植物ゲノムDNAの標的DNA配列に定めるガイドRNA(「gRNA」)と接触させることによる方法であり、編集された植物ゲノムDNAは、反復GGGGS配列以外の第1のリンカーを有する融合タンパク質により編集された植物ゲノムDNAと比較してオフターゲット編集が低い、方法も提供する。一態様では、この融合タンパク質は、配列番号24を含む。
【0010】
本発明者らはまた、オフターゲット編集が低い編集植物の集団を得る方法であって、(a)編集されるゲノムDNAを含む植物細胞の集団を得ること、(b)上記態様の内の1つにより説明されている融合タンパク質及び任意のUGIドメインをコードするヌクレオチド配列を得ること、(c)この植物細胞の集団を工程(b)のヌクレオチド配列で形質転換させ、それにより、この植物細胞の集団内で、この核酸配列によりコードされた融合タンパク質を発現させること、(d)形質転換された植物細胞の集団を植物へと成長させることであり、この植物の少なくとも1つは編集されている、成長させること、並びに(e)工程(d)の生成物から少なくとも1つの編集植物を選択し、それにより編集植物の集団を得ることによる方法であり、この編集植物の集団は、反復GGGGS配列以外の第1のリンカーを有する融合タンパク質により編集された植物と比較してオフターゲット編集が低い、方法も提供する。一態様では、融合タンパク質をコードするヌクレオチド配列は、配列番号17、配列番号24、配列番号35、配列番号39、配列番号43、配列番号50、配列番号52、配列番号54、配列番号56、配列番号81、配列番号83、配列番号85、配列番号87、又は配列番号89を含む。
【図面の簡単な説明】
(【0011】以降は省略されています)
この特許をJ-PlatPat(特許庁公式サイト)で参照する
関連特許
クミアイ化学工業株式会社
多形
11日前
東ソー株式会社
タンパク質の発現方法
1か月前
株式会社トクヤマ
四塩化炭素の製造方法
4か月前
東ソー株式会社
炭素-窒素結合形成方法
4か月前
株式会社トクヤマ
シロキサン類の回収方法
4か月前
株式会社半導体エネルギー研究所
有機化合物
11日前
株式会社半導体エネルギー研究所
有機化合物
2か月前
株式会社トクヤマ
ビオチン誘導体の製造方法
3か月前
栗田工業株式会社
ギ酸の回収方法
1か月前
株式会社トクヤマ
チオラクトン誘導体の製造方法
1か月前
株式会社トクヤマ
ベンザゼピン化合物の製造方法
3か月前
株式会社コスモス
液状炭化水素の増産方法
3か月前
株式会社コスモス
液状炭化水素の増産方法
3か月前
株式会社トクヤマ
ホルムアミド化合物の製造方法
2か月前
東ソー株式会社
イソシアネート化合物の製造方法
2か月前
株式会社トクヤマ
チオファニウム塩誘導体の製造方法
13日前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
3か月前
三菱ケミカル株式会社
アルコールの製造方法
2か月前
国立大学法人東京農工大学
深共晶溶媒
2か月前
本田技研工業株式会社
CO2変換方法
1か月前
大阪瓦斯株式会社
メタン製造システム
1か月前
JNC株式会社
有機ケイ素化合物およびこれを用いた重合体
2か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の製造方法
1か月前
キヤノン株式会社
有機化合物及び有機発光素子
1か月前
株式会社トクヤマ
アシル化ベンゼン誘導体の酸塩の製造方法
2か月前
JNC株式会社
有機ケイ素化合物およびこれを用いた重合体
3か月前
個人
IL-17産生誘導能を有する化合物及びその用途
1か月前
株式会社半導体エネルギー研究所
有機金属錯体、発光デバイス
2か月前
キヤノン株式会社
有機金属錯体及び有機発光素子
1か月前
国立大学法人京都大学
抗がん剤
2か月前
デンカ株式会社
有機酸又はその塩の製造方法
2か月前
岐阜市
化合物、アジュバント組成物および医薬組成物
1か月前
株式会社成和化成
アスコルビン酸誘導体又はその塩及びその化粧料
24日前
株式会社 MTG
ツバキ科ツバキ属の若葉からのエラグ酸製造方法
2か月前
国立大学法人岩手大学
反応性付与化合物
2か月前
三菱ケミカル株式会社
化合物
2か月前
続きを見る
他の特許を見る




 特許ウォッチ
特許ウォッチ