TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025177429
公報種別
公開特許公報(A)
公開日
2025-12-05
出願番号
2024084263
出願日
2024-05-23
発明の名称
原始結節細胞の製造方法
出願人
国立大学法人京都大学
,
株式会社RACTHERA
代理人
弁理士法人京都七条特許事務所
,
個人
,
個人
主分類
C12N
5/0735 20100101AFI20251128BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】椎間板変性症患者への移植適用を実現するために、ヒト多能性幹細胞から中軸中胚葉細胞を含むスフェア状のヒト細胞凝集体をin vitroで効率的に分化誘導する方法及びin vitroで分化誘導された中軸中胚葉細胞を含むスフェア状のヒト細胞凝集体の提供。
【解決手段】以下の特徴を有するスフェア状のヒト細胞凝集体:
(1)原始結節領域(Node area)及び非原始結節領域(Non-Node area)の2つの異なる領域を有し、
(2)原始結節領域及び非原始結節領域は、少なくとも2以上の層構造を有しており、
(3)原始結節領域の表面に存在する第1層に中軸中胚葉細胞が存在し、かつ、原始結節領域には内胚葉細胞は実質的に存在せず、
(4)非原始結節領域の表面に存在する第1層に内胚葉細胞が存在する。
【選択図】なし
特許請求の範囲
【請求項1】
以下の特徴を有するスフェア状のヒト細胞凝集体:
(1)原始結節領域(Node area)及び非原始結節領域(Non-Node area)の2つの異なる領域を有し、
(2)原始結節領域及び非原始結節領域は、少なくとも2以上の層構造を有しており、
(3)原始結節領域の表面に存在する第1層に原始結節細胞が存在し、かつ、原始結節領域には内胚葉細胞は実質的に存在せず、
(4)非原始結節領域の表面に存在する第1層に内胚葉細胞が存在する。
続きを表示(約 720 文字)
【請求項2】
原始結節細胞が、FOXJ1陽性細胞である、請求項1に記載のヒト細胞凝集体。
【請求項3】
FOXJ1陽性細胞が、細胞凝集体全細胞数に対し10%から20%程度の割合で存在する、請求項1又は2に記載のヒト細胞凝集体。
【請求項4】
原始結節細胞が、FOXJ1、BRA及びFOXA2を発現する細胞である、請求項1~3のいずれかに記載のヒト細胞凝集体。
【請求項5】
該細胞凝集体の外側に向かって、原始結節細胞から一次繊毛(primary cilia)が伸びている、請求項1~4のいずれかに記載のヒト細胞凝集体。
【請求項6】
内胚葉細胞がSOX17陽性細胞である、請求項1~5のいずれかに記載のヒト細胞凝集体。
【請求項7】
表面に存在する第1層が、1~10個分の細胞に相当する厚みを有する細胞層から構成される、請求項1~6のいずれかに記載のヒト細胞凝集体。
【請求項8】
非原始結節領域の内側の第2層に中軸中胚葉細胞が存在し、さらにその内側の第3層に胚盤葉上層細胞が存在する、かつ、原始結節領域の内側の第2層に胚盤葉上層細胞が存在する、請求項1~7のいずれかに記載のヒト細胞凝集体。
【請求項9】
胚盤葉上層細胞がOCT4陽性細胞である、請求項8に記載のヒト細胞凝集体。
【請求項10】
原始結節領域及び/又は非原始結節領域において、表面に存在する第1層とその内側に存在する第2層との間に基底膜が存在する、請求項1~9のいずれかに記載のヒト細胞凝集体。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、ヒト多能性幹細胞から中軸中胚葉細胞を含むスフェア状のヒト細胞凝集体をin vitroで効率的に分化誘導する方法及びin vitroで分化誘導された中軸中胚葉細胞を含むスフェア状のヒト細胞凝集体に関する。
続きを表示(約 16,000 文字)
【背景技術】
【0002】
日本では約1300万人が腰痛を患い、その20-40%は椎間板の変性が原因と考えられている。椎間板変性症は、多くの場合において髄核の変性や消失から始まると考えられている。髄核は椎間板の中心部分を構成する組織で、脊椎にクッション性と可動性を与えるが、一度変性した髄核は元に戻ることはない。椎間板変性症を治療する手段として髄核移植による再生治療法の開発が期待されていたが、最近になり、軟骨様髄核をヒトiPS細胞から作製し、生体内の髄核を置換することによって、髄核の機能を補完できることが報告された(非特許文献1)。
【0003】
髄核の分化モデルでは、生体内では胚盤葉上層から原腸陥入を経て、中軸中胚葉に分化し、そこから原始結節及び脊索を経て、髄核に分化すると考えられている。しかし、実際にマウスやヒトES/iPS細胞から髄核細胞の前駆細胞である結節細胞や脊索細胞へ分化を誘導した報告は数報にとどまり(非特許文献2、非特許文献3)、その効率も低く、結節細胞と脊索細胞、又はその前駆細胞である中軸中胚葉への効率的な分化は依然として報告がない。したがって、椎間板変性症患者自身の細胞から髄核を作製し、患者への移植へ適用することは達成されていないままであった。
【先行技術文献】
【非特許文献】
【0004】
Takahashi et al., Biomaterials, 2022, May;284:121491
Loh et al., Cell, 2016, July;14:451-467
Rito et al., bioRxiv, 2023, Feb, 28
【発明の概要】
【発明が解決しようとする課題】
【0005】
従って、本発明の課題は、椎間板変性症患者への移植適用を実現するために、ヒト多能性幹細胞から中軸中胚葉細胞を含むスフェア状のヒト細胞凝集体をin vitroで効率的に分化誘導する方法及びin vitroで分化誘導された中軸中胚葉細胞を含むスフェア状のヒト細胞凝集体を提供することである。
【課題を解決するための手段】
【0006】
本発明者らは、3次元培養技術と最適な培養液を組み合わせることで、hESCs/iPSCから中軸中胚葉細胞を含む凝集体への分化が効率よく誘導され、さらに誘導された中軸中胚葉が生体内で見られるような構造的機能的特性を有することを確認した。さらに本発明者らは、single-cell RNA sequencing(scRNA-seq)解析により、中軸中胚葉細胞を含む凝集体には外胚葉、内胚葉、中胚葉由来の様々な種類の細胞が存在していることを確認し、中軸中胚葉細胞を含む凝集体には中軸中胚葉だけでなく、その周辺領域も含まれており、胚の対応する部分の発生を再現していることを見出した。本発明者らは、これらの知見に基づいてさらに研究を重ねた結果、本発明を完成するに至った。
【0007】
すなわち、本発明は以下の通りのものである。
[1]以下の特徴を有するスフェア状のヒト細胞凝集体:
(1)原始結節領域(Node area)及び非原始結節領域(Non-Node area)の2つの異なる領域を有し、
(2)原始結節領域及び非原始結節領域は、少なくとも2以上の層構造を有しており、
(3)原始結節領域の表面に存在する第1層に原始結節細胞が存在し、かつ、原始結節領域には内胚葉細胞は実質的に存在せず、
(4)非原始結節領域の表面に存在する第1層に内胚葉細胞が存在する。
[2]原始結節細胞が、FOXJ1陽性細胞である、[1]に記載のヒト細胞凝集体。
[3]FOXJ1陽性細胞が、細胞凝集体全細胞数に対し10%から20%程度の割合で存在する、[1]又は[2]に記載のヒト細胞凝集体。
[4]原始結節細胞が、FOXJ1、BRA及びFOXA2を発現する細胞である、[1]~[3]のいずれか一つに記載のヒト細胞凝集体。
[5]該細胞凝集体の外側に向かって、原始結節細胞から一次繊毛(primary cilia)が伸びている、[1]~[4]のいずれか一つに記載のヒト細胞凝集体。
[6]内胚葉細胞がSOX17陽性細胞である、[1]~[5]のいずれかに記載のヒト細胞凝集体。
[7]表面に存在する第1層が、1~10個分の細胞に相当する厚みを有する細胞層から構成される、[1]~[6]のいずれか一つに記載のヒト細胞凝集体。
[8]非原始結節領域の内側の第2層に中軸中胚葉細胞が存在し、さらにその内側の第三層に胚盤葉上層細胞が存在する、かつ、原始結節領域の内側の第2層に胚盤葉上層細胞が存在する、[1]~[7]のいずれか一つに記載のヒト細胞凝集体。
[9]胚盤葉上層細胞がOCT4陽性細胞である、[8]に記載のヒト細胞凝集体。
[10]原始結節領域及び/又は非原始結節領域において、表面に存在する第1層とその内側に存在する第2層との間に基底膜が存在する、[1]~[19]のいずれか一つに記載のヒト細胞凝集体。
[11]非原始結節領域において、基底膜が連続して存在する、[10]に記載のヒト細胞凝集体。
[12]基底膜がLAMININ陽性である、[10]又は[11]に記載のヒト細胞凝集体。
[13]原始結節細胞が、CHORDIN、NOGGIN及びSHHからなる群から選択される1以上の遺伝子を発現する、[1]~[12]のいずれか一つに記載のヒト細胞凝集体。
[13―2]多能性幹細胞由来である、[1]~[13]のいずれか一つに記載のヒト細胞凝集体。
[14](1)分散したヒト多能性幹細胞を迅速に凝集させて、1つの培養コンパートメント中に1つの細胞凝集体を形成させる第一工程、
(2)形成されたヒト多能性幹細胞の細胞凝集体を、基底膜標品を含む培地中で浮遊培養する第二工程、
(3)第二工程で得られた細胞凝集体を、TGFβシグナル伝達経路活性化物質及びWntシグナル伝達経路活性化物質を含む培地中で更に浮遊培養する第三工程、及び
(4)第三工程で得られた細胞凝集体を、Wntシグナル伝達経路活性化物質、BMPシグナル伝達経路阻害物質及びTGFβシグナル伝達経路阻害物質を含む培地中で更に浮遊培養する第四工程を含む、
中軸中胚葉細胞を含むヒト細胞凝集体の製造方法。
[15]前記基底膜標品が、マトリゲルである、[14]に記載の製造方法。
[16]第一工程において、SFEBq法により細胞凝集体を形成させる、[14]又は[15]に記載の製造方法。
[17]第一工程において、Rock阻害物質存在下で細胞凝集体を形成させる、[14]~[16]のいずれか一つに記載の製造方法。
[18]第一工程及び第二工程の培地が、未分化維持因子を含む、[14]~[17]のいずれか一つに記載の製造方法。
[19]第三工程及び第四工程の培地が、未分化維持因子を含む、[18]に記載の製造方法。
[20]第二工程の培養期間が、2日間~4日間である、[14]~[19]のいずれか一つに記載の製造方法。
[21]Wntシグナル伝達経路活性化物質がGSK3阻害物質である、[14]~[20]のいずれかに記載の製造方法。
[22]GSK3阻害物質がCHIR99021である、[21]に記載の製造方法。
[23]TGFβシグナル伝達経路活性化物質がActivin Aである、[14]~[22]のいずれか一つに記載の製造方法。
[24]TGFβシグナル伝達経路阻害物質がALK5阻害物質、及びBMPシグナル伝達経路阻害物質がALK2/3阻害物質である、[14]~[22]のいずれか一つに記載の製造方法。
[25]ALK5阻害物質がSB431542、及びALK2/3阻害物質がLDN-193189である、[24]のに記載の製造方法。
[26]以下の特徴を有するスフェア状のヒト細胞凝集体:
(1)細胞凝集体の最外層にFoxA1陽性の内胚葉細胞が存在する。
(2)細胞凝集体の内側に中軸中胚葉細胞が複数細胞集まって存在し、その周囲に神経管底板及び腹側体節が存在する。
(3)(2)の構造が、1つの細胞凝集体内部に2以上存在する。
[27]神経管底板がNKX6.1及び/又はARXを発現する細胞であり、腹側体節がNkx3.2及び/又はPAX9を発現する細胞である、[26]に記載のヒト細胞凝集体。
[28]最外層が、1個以上10個以下の細胞に相当する厚みを有する細胞層である、[26]又は[27]に記載のヒト細胞凝集体。
[29]細胞凝集体の内側が、最外層から約200μm~300μmの内側である、[26]~[28]のいずれか一つに記載ヒト細胞凝集体。
[30]中軸中胚葉細胞が、BRACHYRY陽性細胞である、[26]~[29]のいずれか一つに記載ヒト細胞凝集体。
[31]細胞凝集体の内側に、中軸中胚葉細胞が5細胞以上50細胞以下集まって存在する、[26]~[30]のいずれか一つに記載ヒト細胞凝集体。
[32]中軸中胚葉細胞が、縦、横及び奥行き各30~40μm程度の範囲内に集まっている、[26]~[31]のいずれか一つに記載ヒト細胞凝集体。
[33]神経管及び体節の数が、それぞれ、1個以上3個以下である、[26]~[32]のいずれか一つに記載ヒト細胞凝集体。
【発明の効果】
【0008】
本発明によれば、ヒト多能性幹細胞から中軸中胚葉細胞を含むスフェア状のヒト細胞凝集体をin vitroで効率的に分化誘導する方法を提供することが可能となる。本発明はまた、中軸中胚葉細胞を含むスフェア状のヒト細胞凝集体を使用することによって、椎間板変性症患者への移植治療を実現することが可能となる。
【図面の簡単な説明】
【0009】
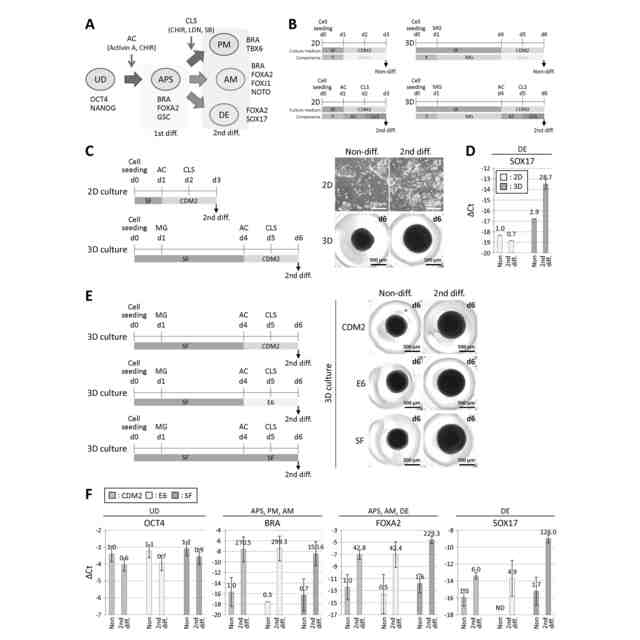
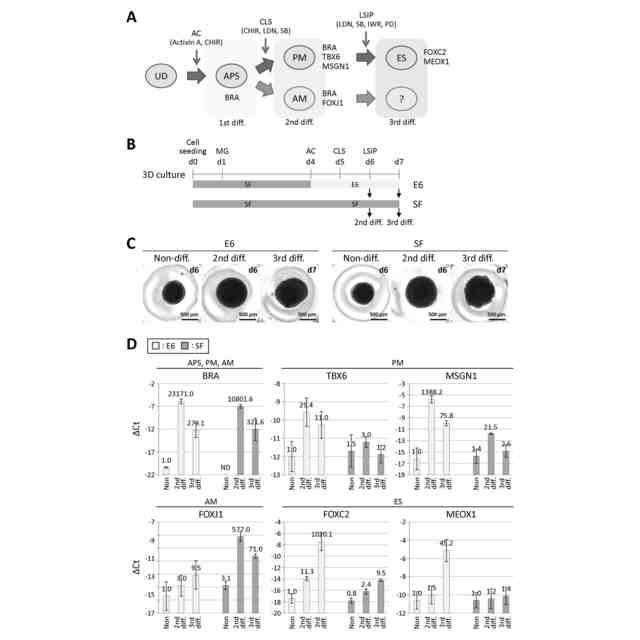
図1は、hESCから中軸中胚葉への分化を示す図である。(A)前方原始線条(APS)から発生する沿軸中胚葉(PM)と中軸中胚葉(AM)の系統分離の予測モデル。中軸中胚葉がAPSから発生するかどうかは、以前は知られていなかった。UD:未分化状態。各系統の代表的なマーカー遺伝子も示した。(B)2D又は3D培養条件下での細胞分化誘導におけるCDM2培地を用いた分化手順の模式図。詳細な手順については図2Bを参照。SF:StemFit培地、CDM2:CDM2培地、AC:Activin A+CHIR、CLS:CHIR+LDN+SB、MG:マトリゲル。(C)第2分化(2nd diff.)を行ったhESCにおける様々な系統マーカー遺伝子のRT-qPCR解析。中軸中胚葉マーカー遺伝子FOXJ1の発現上昇は3次元培養条件下でのみ観察された。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、2次元培養におけるNoneの発現量を1.0としたときの相対発現量(fold change)を示す。UD:未分化状態、APS:前方原始線条、PM:沿軸中胚葉、AM:中軸中胚葉、DE:胚体内胚葉。(D)CDM2、E6、又はSF培地を用いた、3次元培養条件下での細胞分化誘 導中の分化手順の模式図。(E)3次元培養条件下で第2分化を行ったhESCにおけるPMマーカー遺伝子TBX6とAMマーカー遺伝子FOXJ1とNOTOのRT-qPCR解析。SF培地を用いた場合、AMマーカー遺伝子の発現が最も高かった。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を差し引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、CDM2培地で処理したNoneの発現レベルを1.0としたときの相対発現レベル(fold change)を示す。
図2は、中軸中胚葉への分化誘導は胚体内胚葉分化を伴うことを示す図である。(A)胚体内胚葉(DE)は、前方原始線条(APS)からも誘導される。UD:未分化状態。PM:中軸中胚葉。AM:中軸中胚葉。各系統の代表的なマーカー遺伝子を示す。(B)None及び第2分化産物を得るための培養手順、培地、及び使用成分を示す図1Bの詳細模式図。SF:StemFit培地、CDM2:CDM2培地、Y:Y-27632、MG:マトリゲル、AC:ActivinA+CHIR、CLS:CHIR+LDN+SB。(C)図1Bに同じ模式図と、この条件下で培養したhESCの位相差画像。各画像の右上に培養日数を示す。(D)図1B及び図2Cの手順に従って培養したhESCにおけるDEマーカー遺伝子SOX17のRT-qPCR解析。SOX17の発現は、3次元培養の第2分化産物でのみ上昇した。ΔCt値は、β-ACTINのCt値からSOX17のCt値を引くことで算出し、3回実験(n=3)の平均値±標準偏差で示した。各バーの上の数値は、2D培養におけるNoneの発現レベルを1.0としたときの相対発現レベル(fold change)を示す。(E)図1Dに同じ模式図と、これらの条件下で培養したhESCの位相差画像。各画像の右上に培養日数を示す。(F)図1D及び図2Eの手順に従って培養したhESCにおける系統マーカー遺伝子のRT-qPCR解析。SF培地を用いた場合、特に高いDEマーカー遺伝子の発現が観察された。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、CDM2培地で処理したNoneの発現レベルを1.0としたときの相対発現レベル(fold change)を示す。ND:検出されなかった。
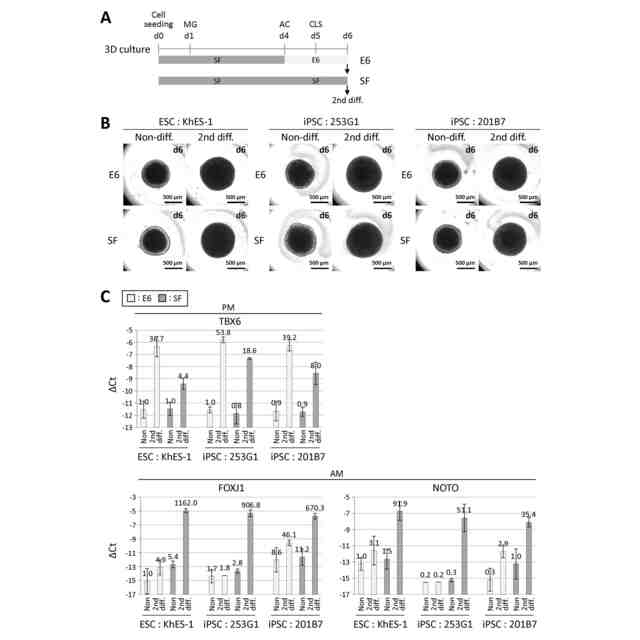
図3は、他の多能性細胞株の中軸中胚葉分化を示す図である。(A)Essential 6(E6)培地又はStemFit(SF)培地を用いた、3次元培養条件下での細胞分化誘導中の分化手順の模式図。MG:マトリゲル、AC:Activin A+CHIR、CLS:CHIR+LDN+SB。(B)(A)の条件で培養した3つのhESC/iPSC株(hESC株:KhES-1、hiPSC株:253G1、hiPSC株:201B7)の位相差画像。各画像の右上に培養日数を示す。(C)3つのhESC/iPSC株における、沿軸中胚葉(PM)マーカー遺伝子TBX6と中軸中胚葉(AM)マーカー遺伝子FOXJ1及びNOTOのRT-qPCR解析。3つの細胞株すべてで同様の遺伝子発現パターンが見られた。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を差し引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、E6培地で処理したKhES-1細胞株におけるNoneの発現レベルを1とした場合の相対発現レベル(fold change)を示す。
図4は、中軸中胚葉分化産物は初期体節にはほとんど分化しないことを示す図である。(A)初期体節(ES)への系統段階モデル。ESはLSIP(LDN、SB、IWR、PD)を用いた3次分化(3rd diff.)により沿軸中胚葉(PM)から誘導されるが、3次分化により中軸中胚葉(AM)から誘導される分化産物は不明であるため、「?」で示す。UD:未分化状態、APS:前方原始線条。(B)3次元培養条件下でEssential 6(E6)培地又はStemFit(SF)培地を用いた3次分化の分化手順の模式図。E6培地を用いた場合、2次分化(2nd diff.)でPMは効果的に誘導されるが、中軸中胚葉(AM)はほとんど誘導されない。一方、SF培地を用いた場合はその逆となる。MG:マトリゲル、AC:ActivinA+CHIR、CLS:chir+ldn+sb、LSIP:LDN+SB+IER+PD。(C)(B)の条件で培養したhESCの位相差画像。各画像の右上に培養日数を示す。(D)(B)と同様の条件で培養したhESCにおける、ESマーカー遺伝子FOXC2とMEOX1、及びその他の系統マーカー遺伝子のRT-qPCR解析。SF培地を用いて細胞を中軸中胚葉に分化させた場合、ESマーカー遺伝子の発現は3回目の拡散後、E6培地を用いた場合よりも有意に低い。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、E6培地で処理したNoneの発現レベルを1.0としたときの相対発現レベル(fold change)を示す。ND:検出されなかった。
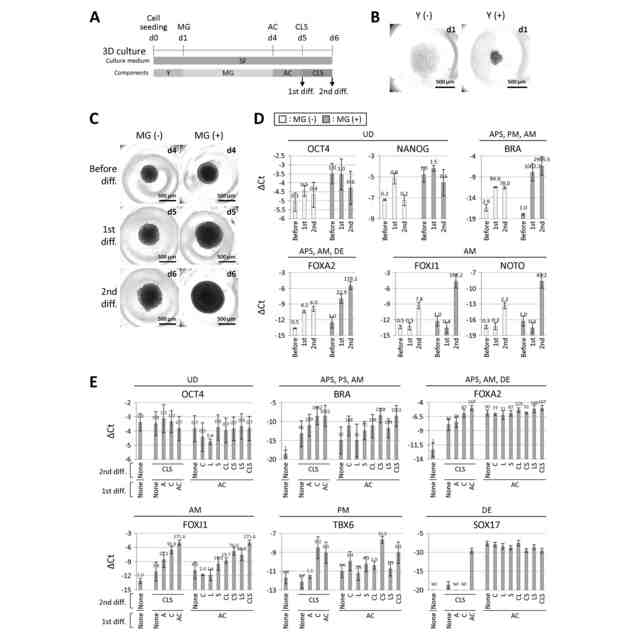
図5は、細胞分化に用いられる各成分の必要性の検討を示す図である。(A)中軸中胚葉分化手順の模式図。培養期間を通してStemFit(SF)培地を使用し、0-1日目はY-27632(Y)、1-4日目はマトリゲル(MG)、4-5日目はActivin A+CHIR(AC)、5-6日目はCHIR+LDN+SB(CLS)を成分とした。(B)0-1日目におけるY-27632処理の必要性の検討。Y(-)はY処理なし、Y(+)はY処理ありの条件で培養したhESCの1日目の位相差画像。Y(-)は細胞死を引き起こし、細胞凝集体を形成しない。各画像の右上に培養日数を示す。(C)1~4日目のマトリゲル(MG)処理の必要性の検討。4日目(分化誘導前[Before diff.])、5日目(1次分化後[1st diff.])、6日目(2次分化後[2nd diff.])のhESCの位相差画像。MG(-)はMG処理なし、MG(+)はMG処理ありの条件下で培養。各画像の右上に培養日数を示す。(D)(C)と同様の条件で培養したhESCで発現した系統マーカー遺伝子のRT-qPCR解析。中軸中胚葉(AM)マーカー遺伝子FOXJ1とNOTOの分化誘導前[Before diff.])、5日目(1次分化後[1st diff.])、6日目(産物における発現は、MG(-)よりもMG(+)の方が高かった。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均±標準偏差で示した。各バーの上の数値は、分化誘導前産物の発現量をMG(+)処理したときの発現量を1.0としたときの相対発現量(fold change)を示す。UD:未分化、APS:前方原始線条、PM:沿軸中胚葉、DE:胚体内胚葉。(E)4-5日目の1次分化におけるActivin AとCHIRの必要性、5-6日目の2次分化におけるCHIR、LDN、IWRI、SBの必要性の検討。各化合物の組み合わせで分化誘導したhESCで発現した系統マーカー遺伝子のRT-qPCR解析。AMマーカー遺伝子FOXJ1の発現が最も高かったのは、細胞をすべての成分で処理した場合であった。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、MG(+)の分化誘導前産物の発現量を1.0としたときの相対発現量(fold change)を示す。ND:検出されなかった。
図6は、中軸中胚葉の分化は、2次元培養よりも3次元培養の方がより効果的に誘導できる。(A)StemFit(SF)培地を用いた中軸中胚葉の分化手順を、2D又は3D培養条件下で培養期間を通して示した模式図。AC:Activin A+CHIR、CLS:CHIR+LDN+SB、MG:マトリゲル。(B)(A)の手順に従い、分化誘導を行わないhESC(なし)、及び1次分化(1st diff.)と2次分化(2nd diff.)を行ったhESCの位相差画像。各画像の右上に培養日数を示す。(C)中軸中胚葉(AM)マーカー遺伝子であるFOXJ1とNOTO、及びその他の系統マーカー遺伝子の、2D又は3D培養hESCにおけるRT-qPCR解析。中軸中胚葉マーカー遺伝子の発現は、2D培養よりも3D培養の方が高いことが観察された。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、2次元培養におけるNoneの発現レベルを1.0とした場合の相対発現レベル(fold change)を示す。(D)3日目又は6日目の2D又は3D培養2次分化産物におけるAMマーカーFOXJ1の免疫染色像。(E)(D)と同じ条件で培養したhESCにおけるFOXJ1陽性領域の割合の定量。DAPI陽性面積に対するFOXJ1陽性面積の比率を計算する。値は4回の実験(n=4)の平均値±標準偏差で示す。
図7は、2D又は3D培養で中軸中胚葉に分化したhESCの免疫染色を示す図である。(A)図6Aに示す手順でクラスター化した未分化(Non-diff.)及び2次分化(2nd diff.)したhESCにおけるFOXJ1(中軸中胚葉マーカー)、BRA(中軸中胚葉、原始線条、汎中胚葉マーカー)、FOXA2(中軸中胚葉、原始線条、中胚葉マーカー)の免疫染色像。2D培養産物は3日目、3D培養産物は分化6日目である。(B)OCT4(未分化上胚葉マーカー)、SOX17(胚体内胚葉マーカー)、FOXA2の免疫染色像。
図7は、2D又は3D培養で中軸中胚葉に分化したhESCの免疫染色を示す図である。(C)FOXJ1、TBX6(沿軸中胚葉マーカー)、及びマウス胚の胚外中胚葉、後方胚盤葉上層、結節に発現するCDX2の免疫染色像。
図8は、1回目(5日目)及び2回目(6日目)産物のシングルセルRNA-seq解析を示す図である。(A)幹細胞マーカーUTF1、原始線条及び汎中胚葉マーカーTBXT(BRAと同じ)、中胚葉マーカーLHX1陽性細胞の1次分化(5日目)におけるUMAP解析。(B)1回目(5日目)と2回目(6日目)の製品を組み合わせたクラスター分析。左パネル:s1:1次分化(5日目)、s2:2次分化、右パネル:遺伝子発現によって分類された11のクラスター。(C)(B)の右パネルに示した各クラスターにおける系統マーカー遺伝子発のヒートマップ。(D)原始線条(PS)、前方原始線条(APS)、中軸中胚葉(AxM)、内胚葉(Endo)、初期沿軸体節中胚葉(沿軸中胚葉と同じ)(PSM)など、系統群を構成する各細胞の位置。(E)UTF1、内胚葉マーカーSOX17、初期体節マーカーFOXC2、中軸中胚葉(原始結節)マーカーNOTOなどの系統マーカー遺伝子を発現する各細胞の位置。(F)擬似時系列解析によって導き出された予想される分化の流れ。未分化の胚盤葉上層細胞はAPS細胞だけでなく、中軸中胚葉前駆細胞(AMP)にも分化する。EndoとPSMはAPSから、AxMはAMPから形成される。(G)各遺伝子の発現レベルの変動を擬似的な時間変化を伴って示すヒートマップ。クラスター1:XXX、クラスター2:YYY、クラスター3:ZZZ。(H)ヒト初期胚のSingle-cell RNA-seq解析との比較(Tyserら、Nature, 2021, Dec;600(7888):285-289)。左のパネル:Tyserの研究のドットが、Tyserの研究と本実施例の結果を組み合わせたクラスタリングのドットに重なっている。右のパネル:Tyserと本実施例の結果を組み合わせたクラスタリングのドットに、本実施例のドットを重ねたもの:内胚葉、Epi:胚盤葉上層、AxM:中軸中胚葉、NE:神経外胚葉、PS:原始線条、SM:体節中胚葉。
図9は、中軸中胚葉に分化した細胞凝集体に見られる原始結節の特徴を示す図である。(A)3次元培養条件下で2次分化(2nd diff.)した6日目のhESC凝集体の免疫染色像。これらの遺伝子発現パターンは、生体内の原始結節における発現パターンを再現している。上図:FOXJ1(中軸中胚葉マーカー)、BRA(中軸中胚葉、原始神経節、汎中胚葉マーカー)、及びFOXA2(軸中胚葉及び中胚葉マーカー)の免疫染色像。下パネル:OCT4(未分化胚盤葉上層マーカー)、SOX17(胚体内胚葉マーカー)、FOXA2の免疫染色。(B)中軸中胚葉分化によって誘導された細胞凝集体の表面における繊毛マーカーAc-α-Tubulin、FOXJ1、SOX17の免疫染色像。灰色の矢印:一次繊毛。(C)中軸中胚葉分化によって誘導された細胞凝集体の走査型電子顕微鏡像。灰色の矢印:一次繊毛。(D)様々な系統に分化したhESCにおける、BMPアンタゴニストである分泌因子遺伝子CHIRDIN、NOGGIN、及び腹側化因子であるソニックヘッジホッグ(SHH)のRT-qPCR解析。これらの遺伝子の発現が最も高かったのは中軸中胚葉(AM)細胞であった。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を差し引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、UD産物の発現量を1.0としたときの相対発現量(fold change)を示す。UD:未分化状態、AM:中軸中胚葉、PM:沿軸中胚葉、DE:胚体内胚葉。
図10は、中軸中胚葉に分化した細胞凝集体における系統マーカー、先端及び基底マーカーの免疫染色解析を示す図である。(A)3次元培養条件下で未分化(Non-diff.)及び2次分化(2nd diff.)した6日目のhESC凝集体の免疫染色像。FOXJ1:中軸中胚葉マーカー、BRA:中軸中胚葉、原始線条、汎中胚葉マーカー、FOXA2:中軸中胚葉、原始線条、中胚葉マーカー、SOX17:胚体内胚葉マーカー、OCT4:未分化胚盤葉上層マーカー、LAMININ:基底マーカー、EZRIN:先端マーカー。(B)(A)の灰色枠で囲んだ非結節部の拡大画像。(C)(A)の灰色の枠で囲まれた結節部の拡大画像。
図11は、中軸中胚葉に分化した細胞凝集体の表面における繊毛マーカータンパク質の発現を示す図である。(A)図12Bの低倍率画像、(B)図12Cの低倍率画像、(C)図8Cの低倍率画像。
図12は、その他の分泌因子遺伝子の発現を示す図である。(A)各系統に分化したhESCにおける各系統マーカー遺伝子のRT-qPCR解析。各系統に分化した細胞は、目的の系統マーカー遺伝子を特異的に発現している。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、UD産物の発現量を1.0としたときの相対発現量(fold change)を示す。UD:未分化状態、AM:中軸中胚葉、PM:沿軸中胚葉、DE:胚体内胚葉。(B)発生初期に発現することが知られている分泌因子遺伝子のRT-qPCR解析。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、UD産物の発現レベルを1.0としたときの相対発現レベル(fold change)を示す。
図13は、中軸中胚葉が神経分化と腹側化を誘導する能力の確認を示す図である。(A)長期の中軸中胚葉分化手順の模式図。6日目、8日目、11日目、14日目はサンプル採取日。SF:StemFit培地、MG:マトリゲル、AC:Activin A+CHIR、CLS:CHIR+LDN+SB。(B)長期の中軸中胚葉分化を行ったhESCにおける中軸中胚葉(AM)マーカー遺伝子FOXJ1とNOTOのRT-qPCR解析。これらの遺伝子の発現は6日目にピークに達し、時間の経過とともに減少した。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、6日目の未分化産物の発現レベルを1.0としたときの相対発現レベル(fold change)を示す。(C)BMPアンタゴニスト遺伝子CHIRDINとNOGGINのRT-qPCR解析。長期の中軸中胚葉分化を行ったhESCにおける腹側化因子遺伝子ソニックヘッジホッグ(SHH)。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、6日目の未分化産物の発現レベルを1.0としたときの相対発現レベル(fold change)を示す。(D)培養上清を用いた神経分化誘導能と腹側化活性の確認実験の模式図。CM_NonとCM_AMはそれぞれ、中軸中胚葉に分化していない細胞凝集体と分化した細胞凝集体の培養上清である。(E)上清で培養したhESCにおける初期神経外胚葉マーカー遺伝子PAX6、SIX2、RXのRT-qPCR解析。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、新鮮なStemFit培地を用いて未分化状態を維持したhESCの発現量を1.0としたときの相対発現量(fold change)を示す。SF+LDNはLDNを添加したSF培地を意味し、その培地で処理したhESCをBMPシグナル阻害による神経分化誘導の陽性対照として用いた。(F)上清で培養したhESCにおけるSHH標的遺伝子HHIPとPTCH1のRT-qPCR解析。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、新鮮なStemFit培地を用いて未分化状態を維持したhESCの発現量を1.0としたときの相対発現量(fold change)を示す。
図14は、長期の中軸中胚葉分化を受けた細胞凝集体の形態と遺伝子発現の変化を示す図である。(A)図13Aと同じ模式図と、この手順に従って培養したhESCの位相差画像。各画像の右上に培養日数を示す。6日目、8日目、11日目、14日目は観察及びサンプル採取のための日数である。SF:StemFit培地、MG:マトリゲル、AC:Activin A+CHIR、CLS:CHIR+LDN+SB(B)長期の中軸中胚葉分化を行ったhESCにおける系統マーカー遺伝子のRT-qPCR解析。全ての遺伝子において、発現は6日目をピークに減少した。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、6日目の未分化産物の発現レベルを1.0としたときの相対発現レベル(fold change)を示す。APS:前方原始線条、PM:沿軸中胚葉、AM:中軸中胚葉、DE:胚体内胚葉。
図15は、中軸中胚葉に分化した細胞凝集体の上清で培養したhESCにおける他の初期神経外胚葉マーカー遺伝子の発現を示す図である。初期神経外胚葉マーカー遺伝子OTX2、ZNF521、及びOCT6のRT-qPCR解析。CM_non及びCM_AMで培養したhESCは、それぞれ非分化及び中軸中胚葉に分化した細胞凝集体の上清である。ΔCt値は、β-ACTINのCt値から各遺伝子のCt値を引いて算出し、3回以上の実験(n≧3)の平均値±標準偏差で示した。各バーの上の数値は、新鮮なStemFit培地を用いて未分化状態を維持したhESCの発現量を1.0としたときの相対発現量(fold change)を示す。SF+LDNはLDNを添加したSF培地を意味し、この培地で処理したhESCをBMPシグナル阻害による神経分化誘導の陽性対照として用いた。
図16は、長期軸中胚葉分化産物のシングルセルRNA-seq解析(11日目)を示す図である。(A)11日目の長期培養細胞凝集体のクラスター解析。遺伝子発現によって9つのクラスターに分類した。(B)TBXTなどの系統マーカー遺伝子を発現する各細胞の位置はBRAと同じ(初期及び後期中軸中胚葉、原始線条、汎中胚葉)、POU5F1(OCT4と同じ)(未分化胚盤葉上層)、FOXJ1(初期中軸中胚葉)、FOXA1(汎内胚葉)、GSC(脊索前板と呼ばれる前方中軸中胚葉)、ARX(神経管の底板)、LBH1(中胚葉)、そしてCHRD(BMPアンタゴニスト)とSHH(腹側化因子)という結節に発現する分泌因子である。(C)FOXC2(初期体節)などの体節関連遺伝子を発現する各細胞の位置、NKX3-2、SOX9、SCX(腹側体節)、SHH標的遺伝子HHIPとPTCH1。
図17は、中軸中胚葉分化を行った長期培養細胞凝集体には、腹側神経管と腹側体節に含まれる細胞が含まれていることを示す図である。(A)長期中軸中胚葉分化手順の模式図。6日目、8日目、11日目、14日目、18日目はサンプル採取日。SF:StemFit培地、MG:マトリゲル、AC:Activin A+CHIR、CLS:CHIR+LDN+SB。(B)後期中軸中胚葉マーカー遺伝子BRAのRT-qPCR解析。値は3回実験(n=3)の平均値±標準偏差で示した。(C)腹側化因子遺伝子SHHのRT-qPCR解析。数値は3回実験(n = 3)の平均値±標準偏差で示した。(D)BRAとSHHの免疫染色像(11日目)。下段は上段の一部拡大。(E)神経管の床板マーカーであるNKX6.1とARXのRT-qPCR解析。数値は3回実験(n=3)の平均値±標準偏差で示した。(F)腹側体節マーカーであるNKX3.2とPAX9のRT-qPCR解析。数値は3回実験(n=3)の平均値±標準偏差で示した。(G)上段と中段:NKX6.1、BRA、SHHの免疫染色像(18日目)。中段は上段の一部を拡大したもの。下段:両底板マーカーNKX6.1とARXの免疫染色像(18日目)。(H) NKX3.2とNKX6.1の免疫染色像(18日目)。(I)8日目からSHHシグナル伝達経路阻害剤シクロパミン(Cyc)で処理した長期培養hESC(14日目)におけるSHH及びSHH標的遺伝子HHIとPTCH1のRT-qPCR解析。数値は3回実験(n=3)の平均値±標準偏差で示した。(J)8日目からシクロパミンで処理した長期培養hESC(14日目)における底板マーカーNKX6.1と蓋板マーカーPAX6のRT-qPCR解析。数値は3回実験(n=3)の平均値±標準偏差で示した。(K)8日目からシクロパミンで処理した長期培養hESC(14日目)の腹側体節マーカーNKX3.2と背側体節マーカーMYODのRT-qPCR解析。V値は3回実験(n=3)の平均値±標準偏差で示した。(L)本実施例で作製した短期(6日目)及び長期(11-18日目)培養細胞凝集体と対応するヒト胚との比較。
図18は、代謝産物中胚葉の長期誘導後に観察された神経外胚葉、体節、腸への分化を示す図である。(A)分化誘導手順の模式図と、分化誘導なし(Non-diff.)と分化した中軸中胚葉(AM diff.)の細胞長凝集体の位相差画像。6、8、11、14、18日目はサンプル採取日。SF:StemFit培地、MG:マトリゲル、AC:Activin A+CHIR、CLS:CHIR+LDN+SB。(B)中脳、中後脳、後脳マーカー遺伝子のRT-qPCR解析。値は平均値±標準偏差で示した。3回以上の実験(n≧3)。(C)体節マーカー遺伝子のRT-qPCR解析。値は平均値±標準偏差で示した。3回以上の実験(n≧3)。(D)汎内胚葉マーカー遺伝子のRT-qPCR解析。値は平均値±標準偏差で示した。3回以上の実験(n≧3)。(E)前腸、後腸、中腸/後腸マーカー遺伝子のRT-qPCR解析。値は平均値±標準偏差で示した。3回以上の実験(n≧3)。(F)一般的な内胚葉マーカーFOXA1(汎内胚葉マーカー、Foxa1)の免疫染色像(14日目)。FOXA1陽性細胞は凝集体表面の細胞層に存在する。(G)基底膜マーカーLAMININの免疫染色像(14日目)。灰色の矢印:BRA陽性脊索様細胞集団を取り囲む基底膜。
図19は、SHHシグナル伝達経路阻害剤シクロパミンの投与は体節の腹側化を阻害することを示す図である。(A)分化誘導とシクロパミン(Cyc)処理手順の模式図と、シクロパミン未処理(-Cyc)と処理(+Cyc)の長細胞凝集体の位相差画像。6、8、11、14、18日目はサンプル採取日。SF:StemFit培地、MG:マトリゲル、AC:Activin A+CHIR、CLS:CHIR+LDN+SB。(B)底板マーカー遺伝子ARXのRT-qPCR解析。(C)腹側体節脳マーカー遺伝子PAX9のRT-qPCR解析。
図20は、中軸中胚葉の系統分離モデルを示す図である。本実施例で示唆された中軸中胚葉(AM)分化の系統分離モデル。未分化の幹細胞は、前方胃形成(APS)以外の経路から発生する前駆細胞(ここでは中軸中胚葉前駆細胞(AMP))を介して中軸中胚葉に分化する。UD:未分化状態、APS:前方胃形成、PM:沿軸中胚葉、DE:胚体内胚葉、AMP:中軸中胚葉前駆細胞、AM:中軸中胚葉。
【発明を実施するための形態】
【0010】
1.本発明の細胞凝集体
本発明は、スフェア状の細胞凝集体(以下、本発明の細胞凝集体)を提供する。本明細書において、細胞凝集体の由来は特に限定されないが、哺乳動物が好ましく、げっ歯類、有蹄類、ネコ目、ウサギ目、霊長類等が包含される。げっ歯類には、マウス、ラット、ハムスター、モルモット等が包含される。有蹄類には、ブタ、ウシ、ヤギ、ウマ、ヒツジ等が包含される。ネコ目には、イヌ、ネコ等が包含される。ウサギ目には、ウサギ等が含包される。霊長類とは、霊長目に属する哺乳動物をいい、霊長類としては、キツネザル、ロリス、ツバイ等の原猿亜目、及びサル、類人猿、ヒト等の真猿亜目が含まれる。
一態様では、本発明の細胞凝集体は、スフィア状のヒト細胞凝集体である。
(【0011】以降は省略されています)
この特許をJ-PlatPat(特許庁公式サイト)で参照する
関連特許
国立大学法人京都大学
抗がん剤
3か月前
国立大学法人京都大学
バイオセンサ
3か月前
国立大学法人京都大学
マグネトロン
6か月前
国立大学法人京都大学
二光子吸収材料
2日前
国立大学法人京都大学
多環芳香族化合物
3か月前
国立大学法人京都大学
多環芳香族化合物
3か月前
国立大学法人京都大学
多環芳香族化合物
2か月前
国立大学法人京都大学
細胞質送達ペプチド
7か月前
国立大学法人京都大学
凍結細胞の製造方法
5か月前
国立大学法人京都大学
高さ調整可能な展示台
6か月前
国立大学法人京都大学
タンパク質集積構造体
1か月前
個人
肝疾患の検出方法
5か月前
国立大学法人京都大学
原始結節細胞の製造方法
1日前
国立大学法人京都大学
剪断流を発生させる器具
8か月前
国立大学法人京都大学
白血病の髄外浸潤マーカー
2か月前
国立大学法人京都大学
近位尿細管生体模倣システム
24日前
国立大学法人京都大学
重合体の製造方法及び重合体
8日前
国立大学法人京都大学
電気伝導体及びその設計方法
3か月前
国立大学法人京都大学
ブロック共重合体の製造方法
5か月前
国立大学法人京都大学
2次元フォトニック結晶レーザ
2か月前
国立大学法人京都大学
陽極およびアルカリ水電解装置
5か月前
国立大学法人京都大学
2次元フォトニック結晶レーザ
2か月前
国立大学法人京都大学
データ読み出し方法および装置
5か月前
国立大学法人京都大学
全固体イオン選択性電極用組成物
5か月前
国立大学法人京都大学
神経変性疾患の予防または治療薬
5か月前
ダイキン工業株式会社
共重合体
24日前
国立大学法人京都大学
配座固定アミロイドβ42誘導体
4か月前
国立大学法人京都大学
老化細胞除去用組成物およびその用途
29日前
国立大学法人京都大学
フーリエ変換装置および無線通信装置
5か月前
国立大学法人京都大学
二酸化炭素を回収する方法および装置
4か月前
DMG森精機株式会社
表面発光装置
3か月前
国立大学法人京都大学
フルオロリン酸イオンを分離する方法
2か月前
国立大学法人京都大学
BRG1遺伝子を標的とした膵癌治療剤
2か月前
住友化学株式会社
組成物及び発光素子
22日前
株式会社構造計画研究所
領域変形装置
4か月前
日本電信電話株式会社
細胞培養デバイス
5か月前
続きを見る
他の特許を見る






 特許ウォッチ
特許ウォッチ