TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025105687
公報種別
公開特許公報(A)
公開日
2025-07-10
出願番号
2025068729,2023099277
出願日
2025-04-18,2017-05-19
発明の名称
ベンゼンスルホンアミド化合物および治療剤としてのそれらの使用
出願人
ゼノン・ファーマシューティカルズ・インコーポレイテッド
,
XENON PHARMACEUTICALS INC.
代理人
個人
,
個人
主分類
C07D
401/12 20060101AFI20250703BHJP(有機化学)
要約
【課題】ベンゼンスルホンアミド化合物および治療剤としてのそれらの使用を提供すること
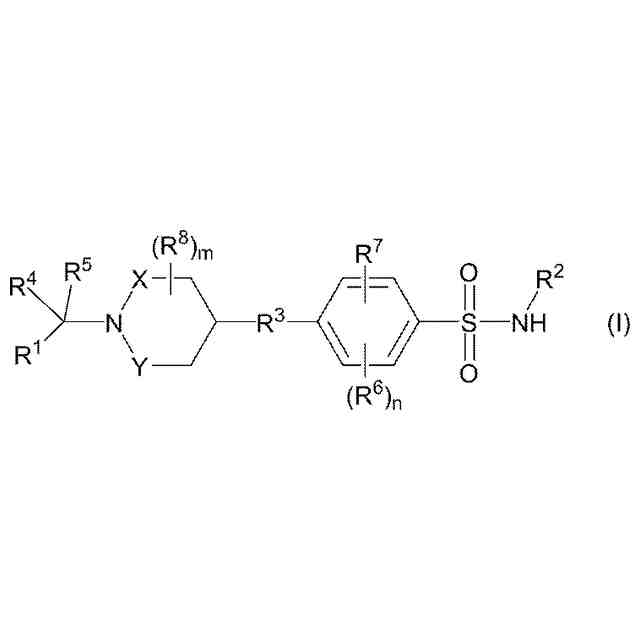
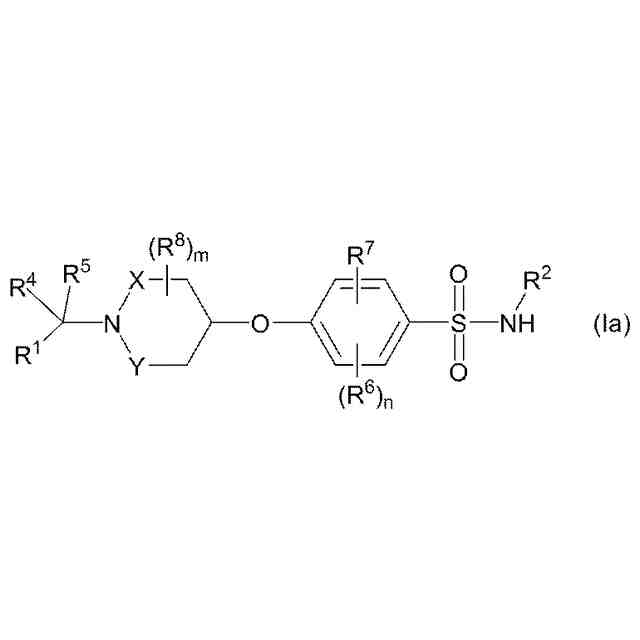
【解決手段】本発明は、電位開口型ナトリウムチャネルに関連する疾患または状態、例えばてんかんの処置のための、ベンゼンスルホンアミド化合物(この化合物は、その立体異性体、エナンチオマー、互変異性体もしくはこれらの混合物としての化合物である)、またはその薬学的に受容可能な塩、溶媒和物もしくはプロドラッグに関する。この実施形態のうちで、好ましい化合物は、4-((1-ベンジルアゼチジン-3-イル)オキシ)-3-クロロ-N-(チアゾール-2-イル)ベンゼン-スルホンアミドである。
【選択図】なし
特許請求の範囲
【請求項1】
てんかん。
発明の詳細な説明
【技術分野】
【0001】
発明の分野
本発明は、ベンゼンスルホンアミド化合物、この化合物を含む薬学的組成物、ならびに、ナトリウムチャネルで媒介される疾患または状態(例えばてんかんおよび/またはてんかん発作障害など)ならびにナトリウムチャネルの媒介に伴う他の疾患および状態の処置において、この化合物およびこの薬学的組成物を使用する方法に関する。
続きを表示(約 5,600 文字)
【背景技術】
【0002】
発明の背景
電位開口型ナトリウムチャネル(Na
V
)は、筋肉および神経における細胞の興奮性の重要な決定因子である(Hille,B,Ion Channels of Excitable Membranes(2001),Sunderland,MA,Sinauer Associates,Inc.)。特に、4つのアイソフォームNa
V
1.1、Na
V
1.2、Na
V
1.3、およびNa
V
1.6は、中枢神経系のニューロンにおけるナトリウム電流の大部分を担う。Na
V
1.3は主として、胚で発現される。新生児期を過ぎると、Na
V
1.1、Na
V
1.2、およびNa
V
1.6が、脳におけるニューロンシグナル伝達を調節する重要なアイソフォームである(Catterall,W.A.,Annual Review of Pharmacology and Toxicology (2014年),第54巻,317-338頁)。
【0003】
Na
V
1.5は、主として心筋細胞(心房、心室、洞房結節、房室結節および心臓のプルキンエ線維が挙げられる)において発現される(Raymond,C.K.ら,J.Biol.Chem.(2004年),第279巻,第44号,46234-41頁)。ヒトNa
V
1.5の変異は、多数の不整脈症候群(例えば、QT3延長(LQT3)、Brugada症候群(BS)、遺伝性心臓伝導欠損、夜間突然死症候群(SUNDS)および乳児突然死症候群(SIDS)が挙げられる)をもたらす(Liu,H.ら,Am.J.Pharmacogenomics(2003年),第3巻,第3号,173-9頁)。ナトリウムチャネル遮断薬治療が、心律動異常を処置する際に広範に使用されている。
【0004】
てんかんとは、脳における興奮性シグナルと抑制性シグナルとの繊細な釣り合いが平衡から外れると起こる、脳における過度の同期の興奮性によって特徴付けられる状態である。これは、過剰な興奮、または抑制の欠乏のいずれかに起因して起こり得る。Na
V
チャネルをコードする遺伝子の変異は、両方の型の不均衡に結び付けられている。
【0005】
Na
V
1.1は、抑制性介在ニューロンの主要なNa
V
アイソフォームとして同定されている(Yu,F.H.ら,Nat.Neurosci.(2006年),第9巻,1142-1149頁)。これらの介在ニューロンは、興奮性グルタミン作用性ニューロンを含めた他の多くのニューロンにシナプスを形成する。介在ニューロンにおける活動電位は、神経伝達物質GABAの、他のニューロンへの放出を誘導し、これらを過分極させ、従って、興奮を抑える。これは、制御されたシグナル伝達を可能にし、局所シグナルが大きい脳領域にわたって広がる興奮の波に拡大することを防ぐ、負のフィードバックをもたらす。抑制性介在ニューロンにおけるこの重要な役割に起因して、Na
V
1.1チャネル機能を損なわせる変異は、これらのニューロンが活性化してGABAを放出することの失敗をもたらし得る(Ogiwara,I.ら,J.Neurosci.(2007年),第27巻,5903-5914頁;Martin,M.S.ら,J.Biol.Chem.(2010年),第285巻,9823-9834頁;Cheah,C.S.ら,Channels(Austin)(2013年),第7巻,468-472頁;およびDut
ton,S.B.,ら,(2013年),第49巻,211-220頁)。その結果は、脳の抑制性トーンの損失、およびグルタミン作用性ニューロンの興奮性の封じ込めの失敗である。この抑制性介在ニューロンの失敗は、脳の領域にわたるニューロンの異常な広範囲規模の同期興奮(てんかん)をもたらし得る。
【0006】
Na
V
1.1をコードする遺伝子(SCN1A)の変異は、2つの広いクラス、すなわち、熱性痙攣プラスを伴う全身てんかん(GEFS+)を引き起こすもの、および乳児重症ミオクロニーてんかん(SMEI)(ドラベ症候群または早期乳児てんかん性脳症6(EIEE6)としても公知)を引き起こすものに入る(McKusik,V.K.ら,A
Epileptic Encephalopathy,Early Infantile 6,EIEE6(2012年),Online Mendelian Inheritance in Man:John Hopkins University)。SMEI変異は、異型接合の常染色体優勢変異であり、そしてしばしば、機能をほとんどまたは全く持たないチャネルをもたらす遺伝子欠失または短縮によって引き起こされる。これらの変異は新規に起こるか、または数例において、無症候性のモザイク親において起こることが示されている(Tuncer,F.N.ら,Epilepsy Research(2015年),第113巻,5-10頁)。患者は、表現型は正常に誕生し、そして痙攣の発症(代表的に、月齢6~年齢1の間)まで、発達のマイルストーンに差し掛かる。この発症の時点は、胚のアイソフォームNa
V
1.3の発現の正常な減少および同時に起こるNa
V
1.1の上昇の結果であると考えられる。Na
V
1.1チャネルが正常レベルに達しない場合、この表現型が明らかになる(Cheah,C.S.ら,Channels(Austin)(2013年),第7巻,468-472頁)。最初の痙攣はしばしば、熱性のエピソードにより誘発され、そしててんかん重積持続状態として現れ得る。痙攣は、人生の最初の数年間にわたって持続し、そして頻度および重篤度が増大し、そして1日あたり100を超えるエピソードの頻度に達し得る。痙攣は、熱によって誘発され得るか、または明白な原因なしに自発的に生じ得る。痙攣の発症後、患者は、発達のマイルストーンを逃し始め、そして重大な認知および行動の不足が生じる(Dravet,C.およびOguni,H.,Handbook of Clinical Neurology(2013年),第111巻,627-633頁)。表現型で診断されたドラベ症候群患者のうちの80~85%が、SCN1A内に原因となる変異を有すると考えられ、一方、患者のうちの他の15~20%は、他の変異を有するか、または、病因が未知である。SMEI患者では、てんかんにおける予期せぬ突然死(SUDEP)が高率で存在し、患者のうち推定37%が、SUDEPによって死亡しているが、この破滅的な転帰に係る機構は不確かなままである(Massey,C.A.,ら,Nature Reviews Neurology(2014年),第10巻,271-282頁)。カルバマゼピンおよびフェニトインのような、電位依存型ナトリウムチャネルを非選択的に標的とする臨床上有用な抗てんかん薬は、SMEI患者に禁忌である。なぜなら、これらの薬は、これらの患者において発作を悪化させ得るからである(Wilmshurst,J.M.ら,Epilepsia(2015年),第56巻,1185-1197頁)。これは、患者がNa
V
1.1機能のさらなる低下に耐えられないせいであろうと推定される。
【0007】
GEFS+はしばしば、比較的穏やかな痙攣表現型と一致する比較的穏やかなチャネル機能不全を誘導する、ミスセンスSCN1A変異によって引き起こされる。多数の、数を増している変異が同定されており、そして表現型の重篤度と浸透度との両方が、かなり変化している。多くのGEFS+患者が痙攣表現型から抜け出すが、全員が抜け出すわけではなく、そして小児期てんかんを有するGEFS+患者は、成人として、一般的な集団よりも、てんかんにかなり罹りやすい。GABA作動性シグナル伝達に関与する他の遺伝子(ナトリウムチャネルの補助的なサブユニットをコードするSCN1B、およびGABA
A
レセプターのサブユニットをコードするGABRG2など)に欠損を引き起こす変異もまた、GEFS+を引き起こし得る(Helbig,I.,Seminars in N
eurology(2015)第35巻,288-292頁)。
【0008】
SMEI患者およびGEFS+患者において同定された同じ変異を抱くトランスジェニックマウスが開発された。両方の場合において、これらのマウスは、ヒト表現型を良好に複製するが、この表現型の浸透性は、遺伝的背景によって有意に影響を受け得る。数個のマウス株は、この変異に対して比較的良好に耐性を有するが、他の下部においては、同じ変異が、深刻な痙攣表現型を引き起こし得る。これらの差異は、興奮性の表現型を調節する他の遺伝子の発現の異なるレベルに起因すると推定される(Miller,A.R.ら,Genes,Brain,and Behavior(2014年),第13巻,163-172頁;Mistry,A.M.ら,Neurobiology of Disease(2014年),第65巻,1-11頁;およびHawkins,N.A.ら,Epilepsy Research(2016年),第119巻,20-23頁)。
【0009】
脳において、Na
V
1.2およびNa
V
1.6は主として、興奮性グルタミン酸作用性ニューロンにおいて発現される。両チャネルは、活動初期セグメント(AIS)(ニューロン細胞体に隣接する、入力を統合するように働き、そしてこの細胞体および遠位の樹状突起への活動電位伝搬を開始させる、ニューロンの領域)において特に濃厚である(Royeck,M.ら,J.Neurophysiol.(2008年),第100巻,2361-2380頁;Vega,A.V.ら,Neurosci.Lett.(2008年),第442巻,69-73頁;およびHu,W.ら,Nat.Neurosci.(2009年),第12巻,996-1002頁)。Na
V
1.6は、初期AIS(細胞体から遠位)に特に濃厚に局在する傾向があり、ここで、活動電位の開始を誘発するように働くと考えられる。Na
V
1.2は、AISの、細胞体に最も近いセグメントにより高度に局在する。SCN2A(Na
V
1.2)とSCN8A(Na
V
1.6)との両方における変異が、てんかんおよび認知遅延に結び付けられている。これらの変異の影響は、チャネル機能に対する影響のレベルと、患者の表現型との両方において、様々である。Na
V
1.2とNa
V
1.6との両方は、末梢ニューロンにおいてもまた発現される。Na
V
1.6は、有髄ニューロンのランヴィエ節において特に濃厚であり、ここで、健康な状態および高速ニューロンシグナル伝達を維持するために重要である。
【0010】
ほんの少数のNa
V
1.2変異のみを記載したが、これらは主として、中枢神経系の病理、特に、てんかんと結び付けられている(Kearney,J.A.ら,Neuroscience(2001年),第102巻,307-317頁;Zerem,A.ら,European Journal of Paediatric Neurology:EJPN:Official Journal of the European Paediatric Neurology Society(2014年),第18巻,567-571頁;Fukasawa,T.ら,Brain & Development(2015年),第37巻,631-634頁;Howell,K.B.ら,Neurology(2015年),第85巻,958-966頁;Saitoh,M.ら,Epilepsy Research(2015年),第117巻,1-6頁;Samanta,D.ら,Acta Neurologica Belgica(2015年),第115巻,773-776頁;Carroll,L.S.ら,Psychiatric Genetics(2016年),第26巻,60-65頁;およびSchwarz,N.ら,Journal of Neurology(2016年),第263巻,334-343頁)。てんかん変異は、本来は、機能変異の増大であると推定される。すなわち、これらの変異はナトリウム電流の量の増大をもたらし、これによって興奮性を増大させる。合理的な疑いの余地なく、インビボでチャネル機能に対する影響を樹立することは困難であり、そしてこれらの変異のうちの数個は依然として、機能表現型の損失をもたらし得る。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
東ソー株式会社
炭素-窒素結合形成方法
13日前
株式会社トクヤマ
グルコンアミド誘導体
3か月前
株式会社トクヤマ
四塩化炭素の製造方法
13日前
株式会社トクヤマ
シロキサン類の回収方法
13日前
日産化学株式会社
ピリジン化合物の製造方法
2か月前
株式会社トクヤマ
ビオチン誘導体の製造方法
1日前
ダイキン工業株式会社
シラン化合物
3か月前
株式会社トクヤマ
グルコンアミド誘導体の製造方法
3か月前
日本特殊陶業株式会社
メタン製造装置
1か月前
ユニチカ株式会社
ビスマレイミドおよびその製造方法
1か月前
artience株式会社
四塩基酸無水物の製造方法
20日前
信越化学工業株式会社
新規化合物
15日前
四国化成工業株式会社
エポキシ化合物およびその利用
2か月前
金剛化学株式会社
ボルチオキセチンの製造方法
1か月前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
23日前
国立大学法人京都大学
細胞質送達ペプチド
3か月前
株式会社トクヤマ
サフィナミド若しくはその塩の製造方法
20日前
四国化成工業株式会社
イソフタル酸化合物およびその利用
2か月前
四国化成工業株式会社
テレフタル酸化合物およびその利用
2か月前
キヤノン株式会社
有機化合物及び有機発光素子
2か月前
株式会社カネカ
プロピレンオキサイド(PO)製造システム
2か月前
ダイキン工業株式会社
SF5含有シラン化合物
3か月前
キヤノン株式会社
有機化合物及び有機発光素子
1か月前
キヤノン株式会社
有機化合物及び有機発光素子
1か月前
旭化成株式会社
トリオキサンの製造方法
23日前
東ソー株式会社
免疫グロブリン結合性タンパク質の保存溶液
20日前
小川香料株式会社
化合物及び香料組成物
3か月前
小川香料株式会社
化合物及び香料組成物
3か月前
キヤノン株式会社
有機化合物及び有機発光素子
1か月前
株式会社半導体エネルギー研究所
有機金属錯体、発光デバイス
1か月前
JFEスチール株式会社
メタノール合成方法
1か月前
旭化成株式会社
ホルムアルデヒドの製造方法
1か月前
株式会社トクヤマ
精製第4級アンモニウム化合物水溶液の製造方法
2か月前
公立大学法人大阪
カルボン酸化合物の製造方法
16日前
株式会社半導体エネルギー研究所
有機化合物、および発光デバイス
27日前
旭化成株式会社
軽質オレフィンの製造方法
1か月前
続きを見る
他の特許を見る
 特許ウォッチ
特許ウォッチ