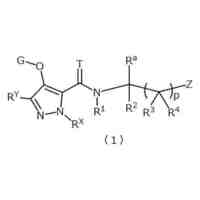
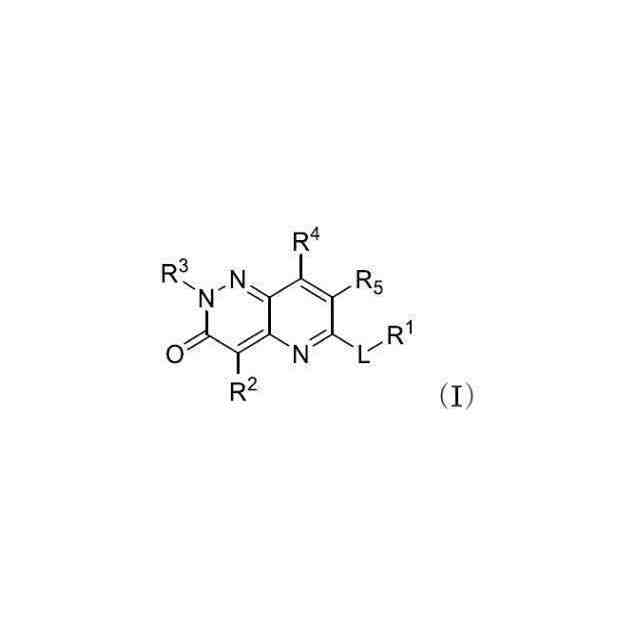
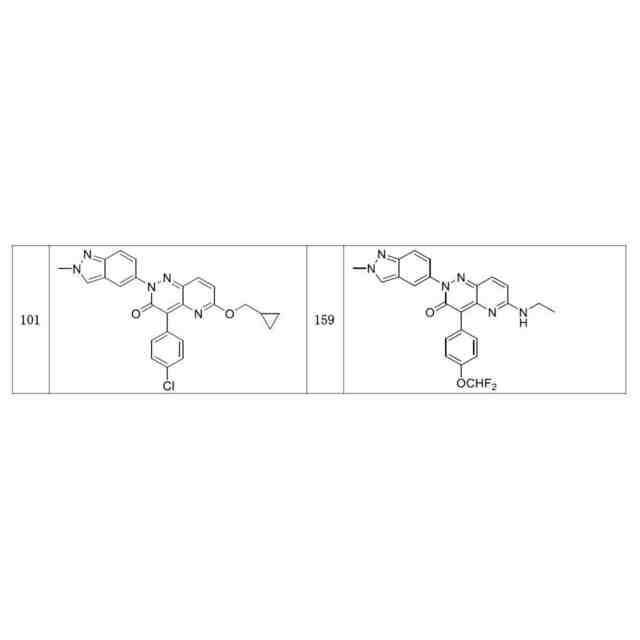
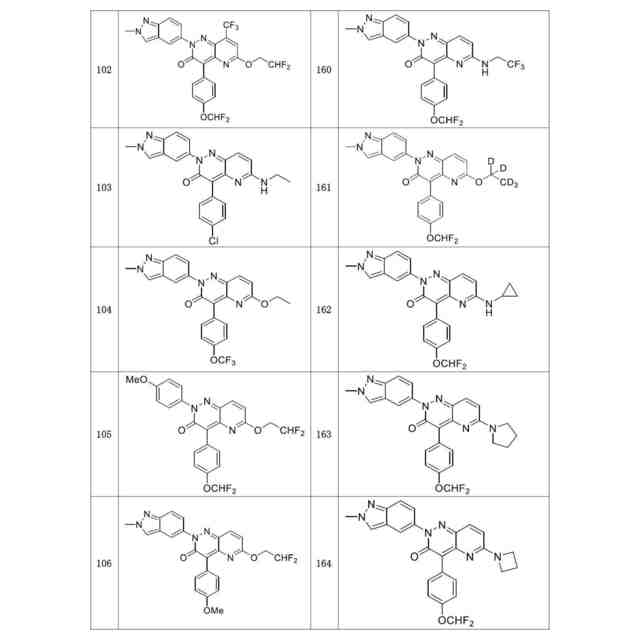
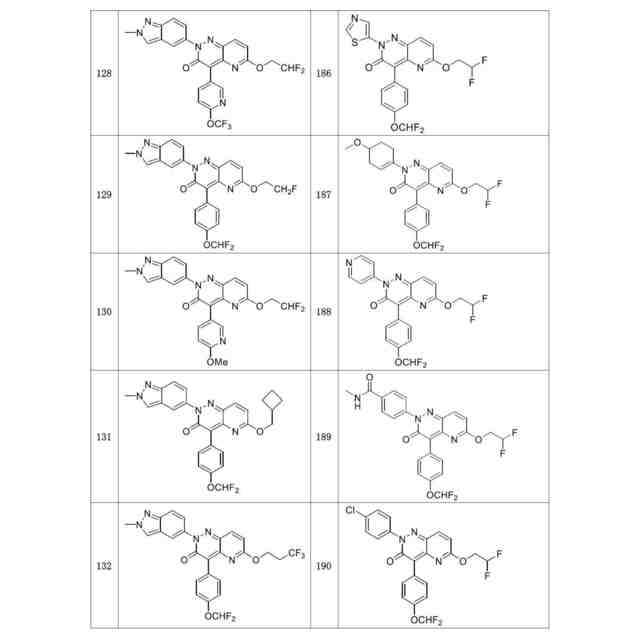
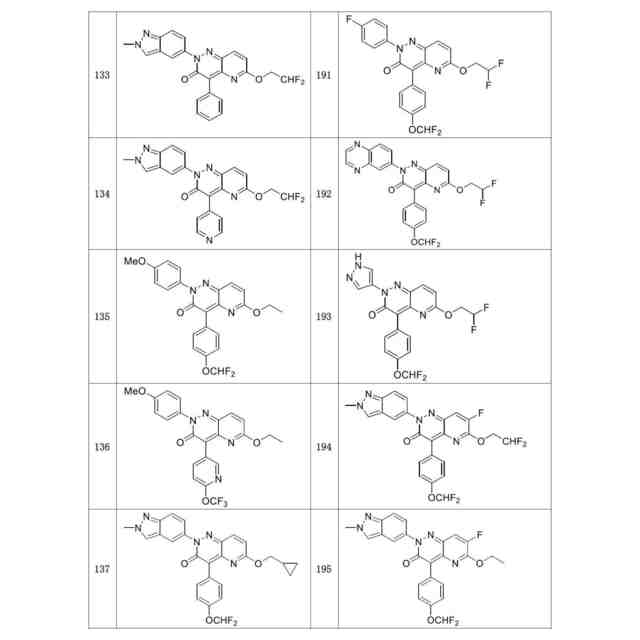
発明の詳細な説明【技術分野】 【0001】 関連出願への相互参照 本出願は、2018年12月27日に出願された米国仮特許出願第62/785,519号の優先権を主張し、その全体が本明細書に参照により組み込まれる。 続きを表示(約 4,000 文字)【背景技術】 【0002】 S-アデノシルメチオニンシンテターゼとしても知られるメチオニンアデノシルトランスフェラーゼ(MAT)は、メチオニンおよびATPからのS-アデノシルメチオニン(SAM、またはADOMET)の合成を触媒する細胞酵素であり、当該触媒はメチオニンサイクルの律速段階であると考えられている。SAMは、ポリアミン生合成のプロピルアミノドナーであり、DNAメチル化の主たるメチルドナーであり、遺伝子転写および細胞増殖に関与し、また二次代謝産物の産生に関与する。 【0003】 MAT1AおよびMAT2Aと表される二つの遺伝子は、それぞれ別個の二つの触媒性MATアイソフォームをコードする。第三の遺伝子、MAT2Bは、MAT2A調節サブユニットをコードする。MAT1Aは成人肝臓で特異的に発現され、一方でMAT2Aは広く分布している。MATアイソフォ ーム同士は触媒運動性および調節性の特性で異なるため、MAT1A発現細胞はMAT2A発現細胞よりも著しく高いSAMレベルを有する。MAT2Aプロモーターの低メチル化およびヒストンアセチル化は、MAT2A発現上昇を引き起こすことが分かっている。例えば、M. Vazquez-Chantada et al., Gastroenterology 138 (2010) 1943-53; M. Frau et al., J. Hepatol. 59 (2013) 830-41; M. Frau et al., Hepatology 56 (2012) 165-75; and R. M. Pascale et al., Transl. Gastroenterol. Hepatol. 3 (2018) 36を参照されたい。 【0004】 肝細胞がん(HCC)では、MAT1Aの発現低下およびMAT2Aの発現上昇が起こるが、これはMAT1A:MAT2Aスイッチとして知られている。このスイッチは、MAT2Bの発現上昇を伴い、より低いSAM含量を導くが、これは肝腫細胞に対する増殖利益を提供する。MAT2Aは、肝腫細胞の増殖を促進する上で重要な役割を果たすため、抗腫瘍療法の標的である。最近の研究では、低分子干渉RNAを使用したサイレンシングが実質的に増殖を抑制し、肝腫細胞にお いてアポトーシスを誘発することが示されている。例えば、T. Li et al., J. Cancer 7(10) (2016) 1317-1327を参照されたい。 【0005】 MTAP欠損である一部のがん細胞株は、特にMAT2Aの阻害に感受性である。Marjon et al. (Cell Reports 15(3) (2016) 574-587)。MTAP(メチルチオアデノシンホスホリラーゼ)は、メチルチオアデノシン(MTA)のアデニンおよび5-メチルチオリボース-1-リン酸塩への変換を触媒する、正常組織で広く発現される酵素である。アデニンは、アデノシン一リン酸を生成するために再利用され、5-メチルチオリボース-1-リン酸塩はメチオニンおよびギ酸塩に変換される。このサルベージ経路のため、MTAは、例えばL-アラノシンなどの代謝拮抗剤を用いて、デノボプリン合成が遮断されたときに代替的プリン源としての役割を果たすことができる。 【0006】 MAT2Aは、肝細胞がんおよび白血病を含むMTAP欠失を欠いているさらなるがんでは調節不全にされる。J. Cai et al., Cancer Res. 58 (1998) 1444-1450; T. S. Jani et al., Cell. Res. 19 (2009) 358-369。RNA干渉を介したMAT2A発現のサイレンシングは、いくつかのがんモデルにおいて抗増殖効果をもたらす。H. Chen et al., Gastroenterology 133 (2007) 207-218; Q. Liu et al. Hepatol. Res. 37 (2007) 376-388。 【0007】 多数のヒトおよびマウスの悪性細胞は、MTAP活性を欠いている。MTAP欠損は組織培養細胞において存在するだけでなく、当該欠損は原発性白血病、神経膠腫、黒色腫、膵がん、非小細胞肺がん(NSCLC)、膀胱がん、星細胞腫、骨肉腫、頭頸部がん、粘液様軟骨肉腫、卵巣がん、子宮内膜がん、乳がん、軟部組織肉腫、非ホジキンリンパ腫、および中皮腫においても存在する。ヒトMTAPをコードする遺伝子は、ヒト染色体9p上の領域9p21に位置する。この領域はまた、腫瘍抑制遺伝子p16INK4A(CDKN2Aとしても知られる)およびpl5INK4Bも含有する。これら遺伝子は、それぞれサイクリンD依存性キナーゼcdk4およびcdk6の阻害剤である、p16およびp15をコードする。 代わりとして、p16INK4A転写物を、pl4ARFをコードする転写物に挿入される代替的なリーディングフレーム(ARF)とすることが可能である。pl4ARFはMDM2に結合し、p53の分解を防止する(Pomerantz et al. (1998) Cell 92:713-723)。9p21染色体領域は、白血病、NSLC、膵がん、神経膠腫、黒色腫、および中皮腫を含む、さまざまながんでホモ接合性欠失していることがよくあるため、関心対象である。欠失は複数の遺伝子を不活性化することが多い。例えば、Cairns et. al. ((1995) Nat. Gen. 11:210-212)は、500を超える原発腫瘍を研究した後に、当該腫瘍において特定されたほぼすべての欠失が 、MTAP、pl4ARF、およびP16INK4Aを含有する170kb領域を含んでいたことを報告した。Carson et al. (国際公開第99/67634号)は、MTAPをコードする遺伝子およびp16をコードする遺伝子の腫瘍発生段階とホモ接合性の喪失との間に相関関係が存在することを報告した。例えば、p16INK4Aではないが、MTAP遺伝子の欠失は、発生の早期段階でのがんの兆候であることが報告されたが、p16およびMTAPをコードする遺伝子の欠失は、腫瘍発生のより進行した段階でのがんの兆候であることが報告された。一部の骨肉腫患者では、MTAP遺伝子は診断時に存在していたが、後の時点では欠失していた(Garcia-Castellano et al., Clin. Cancer Res. 8(3) 2002 782-787)。 【先行技術文献】 【非特許文献】 【0008】 M. Vazquez-Chantada et al., Gastroenterology 138 (2010) 1943-53 M. Frau et al., J. Hepatol. 59 (2013) 830-41 M. Frau et al., Hepatology 56 (2012) 165-75 R. M. Pascale et al., Transl. Gastroenterol. Hepatol. 3 (2018) 36 T. Li et al., J. Cancer 7(10) (2016) 1317-1327 Marjon et al. (Cell Reports 15(3) (2016) 574-587) J. Cai et al., Cancer Res. 58 (1998) 1444-1450 T. S. Jani et al., Cell. Res. 19 (2009) 358-369 H. Chen et al., Gastroenterology 133 (2007) 207-218 Q. Liu et al. Hepatol. Res. 37 (2007) 376-388 Pomerantz et al. (1998) Cell 92:713-723 Garcia-Castellano et al., Clin. Cancer Res. 8(3) 2002 782-787 【発明の概要】 【課題を解決するための手段】 【0009】 本開示は、MAT2Aを阻害する化合物を提供する。化合物およびそれらの医薬組成物は、 例えば外科手術、放射線療法、化学療法、およびホルモン療法などの標準治療に対して不応性のものを含む、種々のがんを治療する方法に有用である。 【0010】 したがって、一部の実施形態によれば、本開示は、以下の一般式Iに従った化合物またはその薬学的に許容可能な塩、互変異性体、および/またはアイソトポログを提供する: JPEG 2025133887000001.jpg 24 47 (【0011】以降は省略されています) この特許をJ-PlatPat(特許庁公式サイト)で参照する

 特許ウォッチ
特許ウォッチ