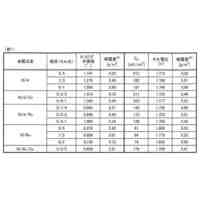
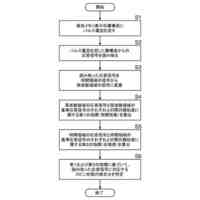
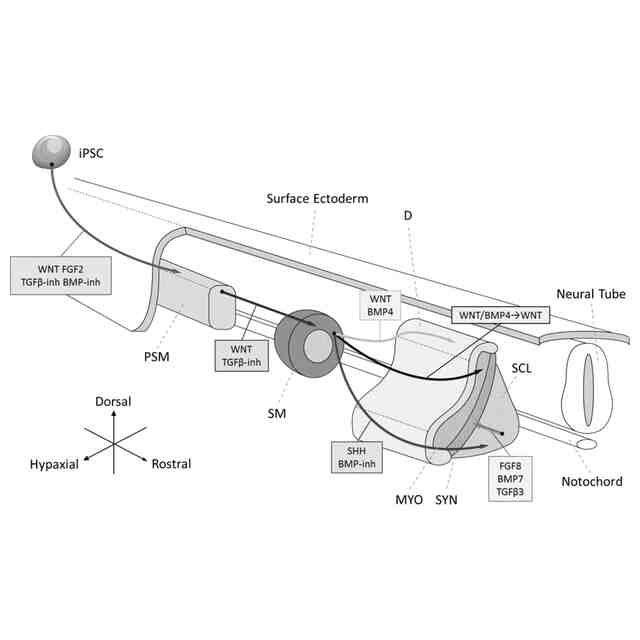
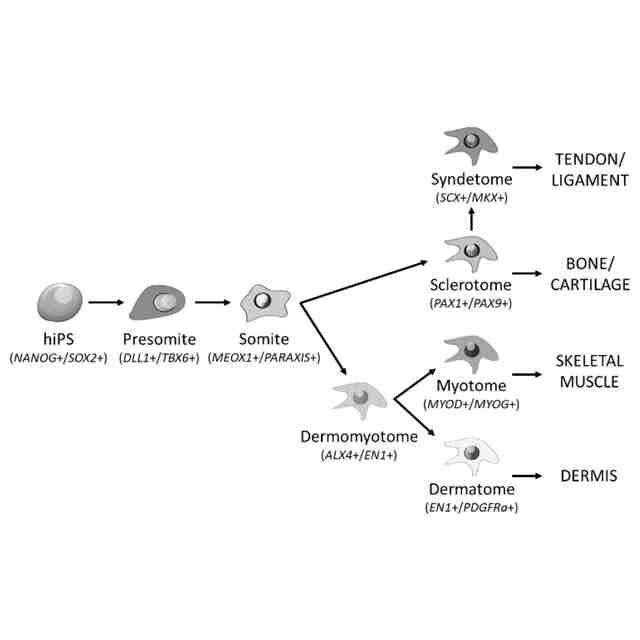
発明の詳細な説明【技術分野】 【0001】 本願は、多能性幹細胞から体節細胞の製造方法に関する。本願はまた、多能性幹細胞から製造された体節細胞を更に分化誘導することにより、多能性幹細胞から筋節細胞、真皮節細胞、硬節細胞、靱帯節細胞および間葉系間質細胞を製造する方法に関する。本願はまた、体節細胞から真皮節細胞、靱帯節細胞および間葉系間質細胞を製造する方法に関する。 続きを表示(約 5,700 文字)【背景技術】 【0002】 内因性シグナル伝達環境の再現は、多能性幹細胞(pluripotent stem cells, PSC)からの所望の細胞種の誘導に重要だと考えられている。発生生物学の知見に基づいて、アクチビン/Nodal/TGFβシグナル伝達が、多能性幹細胞から中内胚葉への分化を誘導し、BMPシグナル伝達が多能性幹細胞から中胚葉への分化を誘導し、これらのシグナルの妨害が神経細胞を誘導することがわかっている(非特許文献1~4)。 【0003】 注目すべきことに、アクチビン/Nodal/TGFβシグナル伝達およびBMPシグナル伝達で誘導された中胚葉は、沿軸中胚葉(神経管と側板中胚葉との間に形成される、中胚葉のサブ集団)ではなく、主に側板中胚葉(中胚葉の側部(腹部)のサブ集団)からなることが報告されている。いくつかの治験が、アクチビン/Nodal/TGFβに基づくプロトコルの改変によって沿軸中胚葉を誘導するために行われたが、誘導の比率は相対的に低い(約20%)ままであったと報告されている(非特許文献5)。 【0004】 近年、いくつかのグループが、異なる考えに基づく沿軸中胚葉の誘導の成功を報告している(非特許文献6~9)。これらの報告では、神経(背側)となる運命へ誘導するため、細胞をアクチビン/Nodal/TGFβを含まない処理、またはTGFβ阻害剤を含む処理を行っており、さらに相対的に高濃度のGSK3阻害剤(WNTシグナル伝達活性化剤)を用いている。これらのプロトコルを用いることで、沿軸中胚葉の誘導率は、約70~95%に達した(非特許文献7および9)。この神経から沿軸中胚葉への変換は、胚形成において神経と沿軸中胚葉に共通の前駆体が存在することを示唆する(非特許文献10および11)。これらの前駆体は神経中胚葉前駆体または軸性中胚葉と呼ばれる。この理論は、Wnt3aノックアウトマウスにおいて、沿軸中胚葉の消失の代わりに異所性(二次)神経管の形成が認められる事実(非特許文献12および非特許文献12)によっても支持される。 【0005】 沿軸中胚葉およびその先の分化の誘導は達成できたが、依然として対処すべきいくつかの問題がある。脊椎動物の発生の過程において、沿軸中胚葉はまず、未分節中胚葉(PSM)を後方に、体節(SM)を前方に形成する。体節は最終的に、背側が皮筋節(DM)に、腹側が硬節に分化する。皮筋節は真皮の前駆体である真皮節(D)および骨格筋の前駆体である筋節(MYO)を生じ、硬節のサブ集団は、腱および靱帯の前駆体である靱帯節を形成する(非特許文献14)。多能性幹細胞から誘導された体節細胞が完全なものであるというためには、誘導された体節細胞が、真皮節細胞、筋節細胞、硬節細胞および靱帯節細胞への分化能を示すことが重要である。上述の報告は、筋節細胞および硬節細胞の誘導には成功しているが、真皮節細胞および靱帯節細胞における誘導プロトコルは、未だ確立されていない。さらに、側板中胚葉は間葉系間質細胞(MSC)の主要な供給源である(非特許文献15)が、体節細胞もまた、間葉系間質細胞の供給源になり得る。しかしながら、多能性幹細胞から、沿軸中胚葉を介して間葉系間質細胞を誘導した報告はない。 【先行技術文献】 【非特許文献】 【0006】 Bernardo, A.S., Faial, T., Gardner, L., Niakan, K.K., Ortmann, D., Senner, C.E., Callery, E.M., Trotter, M.W., Hemberger, M., Smith, J.C., et al. (2011). BRACHYURY and CDX2 mediate BMP-induced differentiation of human and mouse pluripotent stem cells into embryonic and extraembryonic lineages. Cell Stem Cell 9, 144-155. Chambers, S.M., Fasano, C.A., Papapetrou, E.P., Tomishima, M., Sadelain, M., and Studer, L. (2009). Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. Nat Biotechnol 27, 275-280. Fasano, C.A., Chambers, S.M., Lee, G., Tomishima, M.J., and Studer, L. (2010). Efficient derivation of functional floor plate tissue from human embryonic stem cells. Cell Stem Cell 6, 336-347. Sumi, T., Tsuneyoshi, N., Nakatsuji, N., and Suemori, H. (2008). Defining early lineage specification of human embryonic stem cells by the orchestrated balance of canonical Wnt/beta-catenin, Activin/Nodal and BMP signaling. Development 135, 2969-2979. Sakurai, H., Sakaguchi, Y., Shoji, E., Nishino, T., Maki, I., Sakai, H., Hanaoka, K., Kakizuka, A., and Sehara-Fujisawa, A. (2012). In vitro modeling of paraxial mesodermal progenitors derived from induced pluripotent stem cells. PLoS One 7, e47078. Chal, J., Oginuma, M., Al Tanoury, Z., Gobert, B., Sumara, O., Hick, A., Bousson, F., Zidouni, Y., Mursch, C., Moncuquet, P., et al. (2015). Differentiation of pluripotent stem cells to muscle fiber to model Duchenne muscular dystrophy. Nat Biotechnol 33, 962-969. Loh, K.M., Chen, A., Koh, P.W., Deng, T.Z., Sinha, R., Tsai, J.M., Barkal, A.A., Shen, K.Y., Jain, R., Morganti, R.M., et al. (2016). Mapping the Pairwise Choices Leading from Pluripotency to Human Bone, Heart, and Other Mesoderm Cell Types. Cell 166, 451-467. Umeda, K., Zhao, J., Simmons, P., Stanley, E., Elefanty, A., and Nakayama, N. (2012). Human chondrogenic paraxial mesoderm, directed specification and prospective isolation from pluripotent stem cells. Sci Rep 2, 455. Xi, H., Fujiwara, W., Gonzalez, K., Jan, M., Liebscher, S., Van Handel, B., Schenke-Layland, K., and Pyle, A.D. (2017). In Vivo Human Somitogenesis Guides Somite Development from hPSCs. Cell Rep 18, 1573-1585. Gouti, M., Delile, J., Stamataki, D., Wymeersch, F.J., Huang, Y., Kleinjung, J., Wilson, V., and Briscoe, J. (2017). A Gene Regulatory Network Balances Neural and Mesoderm Specification during Vertebrate Trunk Development. Developmental cell 41, 243-261 e247. Takemoto, T., Uchikawa, M., Yoshida, M., Bell, D.M., Lovell-Badge, R., Papaioannou, V.E., and Kondoh, H. (2011). Tbx6-dependent Sox2 regulation determines neural or mesodermal fate in axial stem cells. Nature 470, 394-398. Takada, S., Stark, K.L., Shea, M.J., Vassileva, G., McMahon, J.A., and McMahon, A.P. (1994). Wnt-3a regulates somite and tailbud formation in the mouse embryo. Genes & development 8, 174-189. Yoshikawa, Y., Fujimori, T., McMahon, A.P., and Takada, S. (1997). Evidence that absence of Wnt-3a signaling promotes neuralization instead of paraxial mesoderm development in the mouse. Developmental biology 183, 234-242. Brent, A.E., Schweitzer, R., and Tabin, C.J. (2003). A somitic compartment of tendon progenitors. Cell 113, 235-248. Sheng, G. (2015). The developmental basis of mesenchymal stem/stromal cells (MSCs). BMC developmental biology 15, 44. 【発明の概要】 【発明が解決しようとする課題】 【0007】 本願は多能性幹細胞から体節細胞を製造する方法を提供することを目的とする。本願の別の目的は、体節細胞から真皮節細胞を製造する方法を提供することである。本願の更に別の目的は、体節細胞から靱帯節細胞を誘導する方法を提供することである。本願の更に別の目的は、体節細胞から間葉系間質細胞を誘導する方法を提供することである。 【0008】 本願はまた、多能性幹細胞から体節細胞を分化誘導し、体節細胞をさらに筋節細胞、真皮節細胞、硬節細胞および靱帯節細胞、並びに間葉系間質細胞へそれぞれ分化誘導することにより、多能性幹細胞から筋節細胞、真皮節細胞、硬節細胞、靱帯節細胞および間葉系間質細胞を誘導する方法を提供することを目的とする。 【課題を解決するための手段】 【0009】 本願の方法の全容(但し間葉系間質細胞の製造以外)を図1に示す。多能性幹細胞から、インビトロで未分節中胚葉(PSM)を介して体節細胞(SM)を製造した。得られた体節細胞は、生体において体節から更に分化誘導されることが知られている、真皮節(D)、筋節(MYO)、硬節(SCL)および靱帯節(SYN)それぞれに段階的にインビトロで分化させることができた。各誘導プロトコルを、決定した。なお、この図は、論文(Buckingham et al., 2003)から改変した図である。 【0010】 本願は多能性幹細胞を提供する工程、および 多能性幹細胞を、GSK3β阻害剤を含む培地で培養する工程を含む、多能性幹細胞から体節細胞を製造する方法を提供する。本態様において、多能性幹細胞をGSK3β阻害剤を含む培地で培養する工程は好ましくは、 (1)多能性幹細胞をGSK3β阻害剤を含む培地で培養して未分節中胚葉細胞培養物を得る工程、および (2)未分節中胚葉細胞培養物を、GSK3β阻害剤を含む培地で培養して体節細胞培養物を得る工程を含んでいる。 (【0011】以降は省略されています) この特許をJ-PlatPatで参照する




 特許ウォッチ
特許ウォッチ