TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025113280
公報種別
公開特許公報(A)
公開日
2025-08-01
出願番号
2025080652,2021575441
出願日
2025-05-13,2020-06-19
発明の名称
キメラ抗原受容体設計で実施するためのウルトラモジュラー(ultramodular) IgG3ベーススペーサードメイン及び多機能部位
出願人
ユリウス-マクシミリアン-ウニヴェルシテート・ヴュルツブルク
代理人
個人
,
個人
,
個人
主分類
C07K
19/00 20060101AFI20250725BHJP(有機化学)
要約
【課題】本発明が解決しようとする課題は、キメラ抗原受容体(CAR)改変T細胞等の免疫細胞を使用する免疫療法を提供することである。
【解決手段】本発明は、新規のIgG3ヒンジベーススペーサードメインを有し、標的抗原に対する微細に調節された応答を可能にするキメラ抗原受容体(CAR)改変T細胞を使用する免疫療法に関する。
【選択図】なし
特許請求の範囲
【請求項1】
1つ又は複数のIgG3中央部ヒンジリピートドメインモチーフを含む免疫受容体であって、IgG3 CH2及び/又はCH3ドメインを含まない前記免疫受容体。
続きを表示(約 1,200 文字)
【請求項2】
[A-B
n
]
(式中、
Aは配列番号2のアミノ酸配列であり、
Bは前記IgG3中央部ヒンジドメインリピートモチーフであり、前記モチーフは配列番号1のアミノ酸配列を有し、
nは0、1、2、3、4、5、6、7、8、9、10、11、12、13、14及び15からなる群から選択され、好ましくは1~15の間の整数、より好ましくは1~10の間の整数、更により好ましくは1~5の間の整数、最も好ましくは3~5の間の整数である)
のアミノ酸配列に対して少なくとも80%の配列同一性、好ましくは少なくとも90%の配列同一性、又は最も好ましくは100%の配列同一性を有するアミノ酸配列を含む、請求項1に記載の免疫受容体。
【請求項3】
[A-B
n
]のアミノ酸配列と100%の配列同一性を有するアミノ酸配列を含む、請求項2に記載の免疫受容体。
【請求項4】
nが1~10の間の整数である、請求項2又は3に記載の免疫受容体。
【請求項5】
nが1~5の間の整数である、請求項2又は3に記載の免疫受容体。
【請求項6】
nが3~5の間の整数である、請求項2又は3に記載の免疫受容体。
【請求項7】
細胞外抗原結合ドメインと、
スペーサードメインと、
膜貫通ドメインと、
細胞内シグナル伝達ドメインと
を含み、
スペーサードメインが、細胞外抗原結合ドメインと膜貫通ドメインの間に位置し、
場合によりスペーサードメインが、1つ又は複数のIgG3中央部ヒンジドメインリピートモチーフを含む、請求項1から6のいずれか一項に記載の免疫受容体。
【請求項8】
膜貫通ドメイン及び細胞内ドメインが共に、配列番号109、110、111、112、113、114、115及び174からなる群から選択される配列からなる、請求項7に記載の免疫受容体。
【請求項9】
第1のドメインと、
リンカーと、場合により、
第2のドメインと
を含む細胞外抗原結合ドメインを含み、
場合により、リンカーが第1のドメインと第2のドメインの間に位置し、
場合によりリンカーが1つ又は複数のIgG3中央部ヒンジドメインリピートモチーフを含む、請求項1から8のいずれか一項に記載の免疫受容体。
【請求項10】
スペーサードメインが、1つ若しくは複数のIgG3中央部ヒンジドメインリピートモチーフを含み、
及び/又は
細胞外抗原結合ドメインに含まれるリンカーが、1つ若しくは複数のIgG3中央部ヒンジドメインリピートモチーフを含む、
請求項7、8又は9に記載の免疫受容体。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は概して、キメラ抗原受容体(CAR)改変T細胞等の免疫細胞を使用する免疫療法に関する。特に、本発明は、新規のIgG3ヒンジベーススペーサードメインを有し、標的抗原に対する微細に調節された応答を可能にする、キメラ抗原受容体(CAR)改変T細胞を使用する免疫療法に関する。更に、本発明は、1つ又は複数のIgG3ヒンジベースの多機能部位(MFS)をCAR及び他の免疫受容体に導入し、CAR T細胞の精製、刺激、拡大及び枯渇を可能にすることに関する。本発明は、このモチーフを標的にし、前述の機能の遂行を可能にする抗体の配列も含む。
続きを表示(約 18,000 文字)
【背景技術】
【0002】
キメラ抗原受容体(CAR)は、腫瘍細胞の表面抗原を認識するようにT細胞に再指示することを意図して開発された合成免疫受容体である。そのほとんどの基本フォーマットでは、CARは、膜貫通ドメイン及びCD3ζのシグナル伝達ドメインに融合されたモノクローナル抗体の可変重鎖及び可変軽鎖を(シスで、すなわち一本鎖可変断片、scFvとして)含む
1
。この基本CAR設計を改善する方法は、CARによる抗原結合を促進するために範囲及び柔軟性を提供するようにscFvと膜貫通ドメインの間に位置するスペーサードメインを含めることであった
2
。結局、いくつかのスペーサードメインが、IgG1及びIgG4、IgD、CD4、CD7、CD8α及びCD28に由来するFc領域及び免疫グロブリン様ドメインを含むCAR構築物において使用された
3-6
。
【0003】
本分野における従来のアプローチは、たとえ所与の抗原の異なるエピトープ、又は異なる抗原を認識できるとしても、全てのCAR構築物に単一スペーサー設計を使用することである(「1つのCARが全てに適合しなければならない」)。しかし、CARは腫瘍細胞の表面抗原に結合するため、最適な抗原結合、及びCAR修飾T細胞と腫瘍細胞の間の最適な相互作用を可能にする空間要件は、エピトープ及び標的抗原に応じて異なり得る。したがって、全てのエピトープ及び抗原に対して単一スペーサー設計を使用する従来のアプローチは、単純で最適以下のように思われる。最適以下のCAR結合及び/又はCAR修飾T細胞と腫瘍細胞の間の最適以下の相互作用があるならば、続いて起こるCAR-T細胞刺激及び抗腫瘍応答も最適以下になる可能性がある
3、7
。より最適化されたCAR標的分子相互作用を達成する機会を増やすために、本発明者らは、長さ及び組成が異なるIgG4由来スペーサーのバリアントを以前に研究した。浮上したパラダイムは、スペーサー長と有効性の間には相関関係があり、それにより標的細胞の膜近位エピトープはより長いスペーサーを含有するCARによってより良好に到達され、膜遠位エピトープはより短いスペーサーを含有するCARによってより良好に到達されるということであった
7
。IgG4分子の構造に基づき、>100aaずつサイズが異なる3つのIgG4ヒンジベーススペーサーバリアントが利用可能である(IgG4_短: IgG4ヒンジ、12aa; IgG4_中央部: IgG4ヒンジ+C
H
3、119aa; IgG4_長: IgG4ヒンジ+CH
2
+CH
3
、228aa)
7
。
【0004】
全てのヒトIgG分子のうち、IgG3は、最も高いFab-Fab折り畳み柔軟性及びFab Fc折り畳み柔軟性を示す。全ての他の免疫グロブリンとは対照的に、IgG3のヒンジとしてのIgG3の構造は独特であり、エクソン重複に起因する15aaモチーフの3コピーを組み込んでいる
8-11
。ヒンジ領域中にこのモチーフの1又は2コピーのみを有するIgG3の天然に存在するバリアントは、はるかに短いFabとFcの距離を示す(105Åと比べて45Å及び65Å)
8
。これらの段階的違い、並びにこの15aaモチーフの1つ又は複数のコピーの付加又は除去によってスペーサー領域を延長及び短縮する機会により、本発明は、スペーサーの長さを標的ごとに最適設定に微調整し得ることを利用してIgG3ヒンジライブラリーを構築するに至った。更に、本発明者らは、IgG3中央部ヒンジモチーフに特異的なモノクローナル抗体を同定し、CAR T細胞の精製、刺激、拡大及び枯渇を含む、抗原非依存性であるがCAR特異的な追加の機能の利用を可能にした。
【発明の概要】
【課題を解決するための手段】
【0005】
本発明は概して、キメラ抗原受容体(CAR)改変T細胞等の免疫細胞を使用する免疫療法に関する。特に、本発明は、新規のIgG3ヒンジベーススペーサードメインを有し、標的抗原に対する微細に調節された応答を可能にするキメラ抗原受容体(CAR)改変T細胞を使用する免疫療法に関する。更に、本発明は、1つ又は複数のIgG3ヒンジベースの多機能部位(MFS)をCAR及び他の免疫受容体に導入し、CAR T細胞の精製、刺激、拡大及び枯渇を可能にすることに関する。本発明は、このモチーフを標的にし、前述の機能の遂行を可能にする抗体の配列も含む。
【0006】
本発明は、特に以下の項目を提供し、以下の項目を特徴とする。
1. 1つ又は複数のIgG3中央部ヒンジリピートドメインモチーフを含む免疫受容体であって、IgG3 CH2及び/又はCH3ドメインを含まない前記免疫受容体。
2. [A-B
n
]
(式中、
Aは配列番号2のアミノ酸配列であり、
Bは前記IgG3中央部ヒンジドメインリピートモチーフであり、前記モチーフは配列番号1のアミノ酸配列を有し、
nは0、1、2、3、4、5、6、7、8、9、10、11、12、13、14及び15からなる群から選択され、好ましくは1~15の間の整数、より好ましくは1~10の間の整数、更により好ましくは1~5の間の整数、最も好ましくは3~5の間の整数である)
のアミノ酸配列と少なくとも80%の配列同一性、好ましくは少なくとも90%の配列同一性、又は最も好ましくは100%の配列同一性を有するアミノ酸配列を含む、項目1に記載の免疫受容体。
3. [A-B
n
]のアミノ酸配列を有する100%の配列同一性を有するアミノ酸配列を含む、項目2に記載の免疫受容体。
4. nが1~10の間の整数である、項目2又は3に記載の免疫受容体。
5. nが1~5の間の整数である、項目2又は3に記載の免疫受容体。
6. nが3~5の間の整数である、項目2又は3に記載の免疫受容体。
7.細胞外抗原結合ドメインと、
スペーサードメインと、
膜貫通ドメインと、
細胞内シグナル伝達ドメインと
を含み、スペーサードメインが、細胞外抗原結合ドメインと膜貫通ドメインの間に位置し、
場合によりスペーサードメインが、1つ又は複数のIgG3中央部ヒンジドメインリピートモチーフを含む、項目1から6のいずれか一項に記載の免疫受容体。
8.膜貫通ドメイン及び細胞内ドメインが共に、配列番号109、110、111、112、113、114、115及び174からなる群から選択される配列からなる、項目7に記載の免疫受容体。
9.第1のドメインと、
リンカーと、場合により、
第2のドメインと
を含む細胞外抗原結合ドメインを含み、
場合により、リンカーが第1のドメインと第2のドメインの間に位置し、
場合によりリンカーが、1つ又は複数のIgG3中央部ヒンジドメインリピートモチーフを含む、項目1から8のいずれか一項に記載の免疫受容体。
10.スペーサードメインが、1つ若しくは複数のIgG3中央部ヒンジドメインリピートモチーフを含み、
及び/又は
細胞外抗原結合ドメインに含まれるリンカーが、1つ若しくは複数のIgG3中央部ヒンジドメインリピートモチーフを含む、
項目7、8又は9に記載の免疫受容体。
11.T細胞受容体(TCR)、好ましくは組換えTCR; B細胞受容体(BCR)、好ましくは組換えBCR;及びキメラ抗原受容体(CAR)からなる群から選択される、項目1から10のいずれか一項に記載の免疫受容体。
12.免疫受容体が抗原結合ドメインを含み、
I)第1のドメインが重鎖可変ドメインを含む、
II)第1のドメインが軽鎖可変ドメインを含む、
III)第1のドメインが重鎖可変ドメインを含み、第2のドメインが軽鎖可変ドメインを含む、
IV)第1のドメインが重鎖可変ドメインを含み、第2のドメインが重鎖可変ドメインを含む、又は
V)第1のドメインが軽鎖可変ドメインを含み、及び第2のドメインが軽鎖可変ドメインを含む、項目9から11のいずれか一項に記載の免疫受容体。
13.免疫受容体が抗原結合ドメインを含み、前記抗原結合ドメインが、一本鎖可変断片(scFv)の一部である第1のドメイン、リンカー、及び第2のドメインを含み、
scFvが場合により、第1/第2のドメインに含まれる重鎖/軽鎖可変配列として、以下の抗原:
A) CD19、場合により、
i)重鎖可変ドメインは、配列番号27に対して少なくとも80%の配列同一性、好ましくは少なくとも90%の配列同一性を有するアミノ酸配列を有し、
軽鎖可変ドメインは、配列番号28に対して少なくとも80%の配列同一性、好ましくは少なくとも90%の配列同一性を有するアミノ酸配列を有し、
scFvは、CD19に特異的に結合することができる、又は
ii)重鎖可変ドメインは配列番号27のアミノ酸配列を有し、軽鎖可変ドメインは配列番号28のアミノ酸配列を有する;
B) CD20、場合により、
i)重鎖可変ドメインは、配列番号30に対して少なくとも80%の配列同一性、好ましくは少なくとも90%の配列同一性を有するアミノ酸配列を有し、
軽鎖可変ドメインは、配列番号29に対して少なくとも80%の配列同一性、好ましくは少なくとも90%の配列同一性を有するアミノ酸配列を有し、
scFvは、CD20に特異的に結合することができる、又は
ii)重鎖可変ドメインは配列番号30のアミノ酸配列を有し、軽鎖可変ドメインは配列番号29のアミノ酸配列を有する;
【図面の簡単な説明】
【0007】
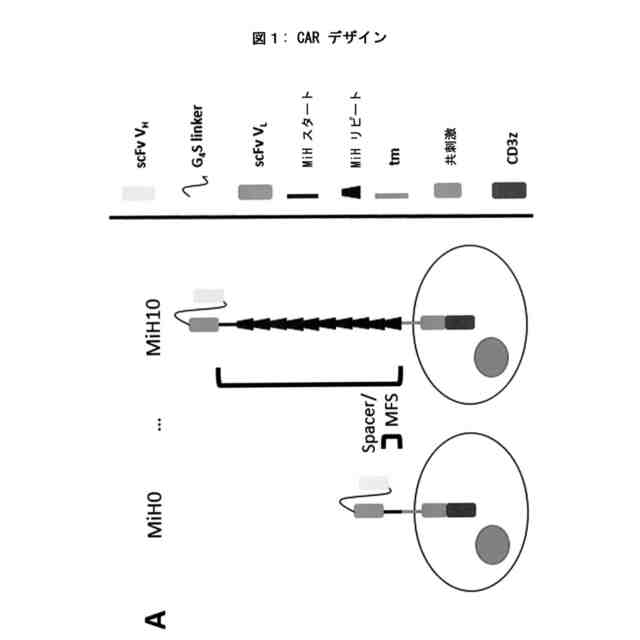
CARの設計。A. 異なる長さのIgG3由来スペーサーを有するCARの概略説明図。
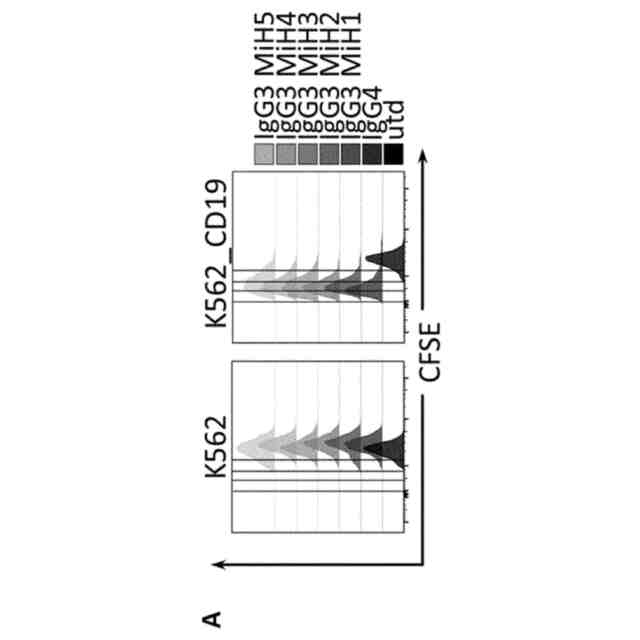
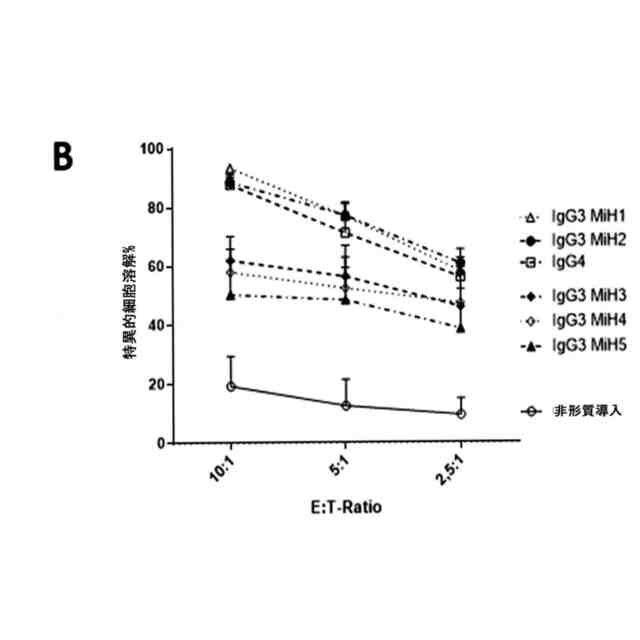
IgG3由来スペーサーを備えるCD19 CAR T細胞はin vitroで機能的である。A. 抗原特異的増殖。CD8
+
T細胞をCFSEで標識し、CD19の発現を伴う又は伴わない照射したK562腫瘍細胞を用いて4:1のE:T比で刺激した。CFSEの希釈によって可視化した増殖を、72時間後に判定した。外因性サイトカインは添加しなかった。n=3の独立した実験についての代表的な例。B. 生物発光系アッセイ(5時間のインキュベーション)における、CD19
+
Jeko-1細胞に対する様々なスペーサードメインを備えるCD8
+
CD19-CAR T細胞及び形質導入していないT細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=3の独立した実験からの平均±SEMとして表す。C. CD8
+
CD19 CAR T細胞と、CD19の発現を伴う又は伴わないK562標的細胞との24時間の共培養から得られた上清中のIFNγを検出するELISA。T細胞(50,000/ウェル)及び標的細胞(12,500/ウェル)を、三重測定ウェルに播種した。値はn=3の独立した実験からの平均±SEMとして表す。
IgG3由来スペーサーを備えるROR1 CAR T細胞はin vitroで機能的である。scFv 4-2に基づくROR1特異的CARのin vitro機能の評価。A. 抗原特異的増殖。CD8
+
T細胞をCFSEで標識し、ROR1の発現を伴う又は伴わない照射したK562腫瘍細胞(K562_ROR1)を用いて4:1のE:T比で刺激した。CFSEの希釈によって可視化した増殖を、72時間後に判定した。外因性サイトカインは添加しなかった。n=3の独立した実験についての代表的な例。B. 生物発光系アッセイ(5時間のインキュベーション)における、K562_ROR1に対する様々なスペーサードメインを備えるCD8
+
ROR1-CAR T細胞及び形質導入していないT細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=3の独立した実験からの平均±SEMとして表す。C. CD8
+
ROR1 CAR T細胞と、ROR1の発現を伴う又は伴わないK562標的細胞との24時間の共培養から得られた上清中のIFNγを検出するELISA。T細胞(50,000/ウェル)及び標的細胞(12,500/ウェル)を、三重測定ウェルに播種した。値はn=3の独立した実験からの平均±SEMとして表す。
IgG3由来スペーサーを備えるROR1 CAR T細胞はin vitroで機能的である。scFv R11に基づくROR1特異的CARのin vitro機能の評価。A. 抗原特異的増殖。CD8
+
T細胞をCFSEで標識し、ROR1の発現を伴う若しくは伴わない照射したK562腫瘍細胞(K562_ROR1)又はR11エピトープと腫瘍細胞膜との距離を増大させたROR1の改変されたバージョン(K562_ROR1/E3AK)を用いて4:1のE:T比で刺激した。CFSEの希釈によって可視化した増殖を、72時間後に判定した。外因性サイトカインは添加しなかった。n=3の独立した実験についての代表的な例。B. 生物発光系アッセイ(5時間のインキュベーション)における、K562_ROR1 (B)に対する様々なスペーサードメインを備えるCD8
+
ROR1-CAR T細胞及び形質導入していないT細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=3の独立した実験からの平均±SEMとして表す。C. CD8
+
ROR1 CAR T細胞と、K562_ROR1(C)との24時間の共培養から得られた上清中のIFNγを検出するELISA。T細胞(50,000/ウェル)及び標的細胞(12,500/ウェル)を、三重測定ウェルに播種した。値はn=3の独立した実験からの平均±SEMとして表す。D. ROR1におけるR11エピトープがE3AKリンカーを用いて腫瘍細胞膜からどのように遠くに移動させられるかを説明する概略図。E. 抗原特異的増殖。CD8
+
T細胞をCFSEで標識し、ROR1の発現を伴う若しくは伴わない照射したK562腫瘍細胞(K562_ROR1)又はR11エピトープと腫瘍細胞膜との距離を増大させたROR1の改変されたバージョン(K562_ROR1/E3AK)を用いて4:1のE:T比で刺激した。CFSEの希釈によって可視化した増殖を、72時間後に判定した。外因性サイトカインは添加しなかった。n=3の独立した実験についての代表的な例。F. 生物発光系アッセイ(5時間のインキュベーション)における、K562_ROR1/E3AK (F)に対する様々なスペーサードメインを備えるCD8
+
ROR1-CAR T細胞及び形質導入していないT細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=3の独立した実験からの平均±SEMとして表す。G. CD8
+
ROR1 CAR T細胞と、K562_ROR1/E3AK (G)との24時間の共培養から得られた上清中のIFNγを検出するELISA。T細胞(50,000/ウェル) 及び標的細胞(12,500/ウェル)を、三重測定ウェルに播種した。値はn=3の独立した実験からの平均±SEMとして表す。
IgG3由来スペーサーを備えるCD20 CAR T細胞はin vitroで機能的である。A. 抗原特異的増殖。CD8
+
T細胞をCFSEで標識し、CD20の発現を伴う又は伴わない照射したK562腫瘍細胞を用いて4:1のE:T比で刺激した。CFSEの希釈によって可視化した増殖を、72時間後に判定した。外因性サイトカインは添加しなかった。n=3の独立した実験についての代表的な例。B. 生物発光系アッセイ(3時間のインキュベーション)における、CD20
+
Raji細胞に対する様々なスペーサードメインを備えるCD8
+
CD20-CAR T細胞及び形質導入していないT細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=3の独立した実験からの平均±SEMとして表す。C. CD8
+
CD20 CAR T細胞と、CD20の発現を伴う又は伴わないK562標的細胞との24時間の共培養から得られた上清中のIFNγを検出するELISA。T細胞(50,000/ウェル)及び標的細胞(12,500/ウェル)を、三重測定ウェルに播種した。値はn=2の独立した実験からの平均±SEMとして表す。
IgG3由来スペーサーを備えるSLAMF7 CAR T細胞はin vitroで機能的である。A. 抗原特異的増殖。CD4
+
T細胞をCFSEで標識し、SLAMF7の発現を伴う又は伴わない照射したK562腫瘍細胞を用いて4:1のE:T比で刺激した。CFSEの希釈によって可視化した増殖を、72時間後に判定した。外因性サイトカインは添加しなかった。n=3の独立した実験についての代表的な例。B. 生物発光系アッセイ(3時間のインキュベーション)における、SLAMF7
+
MM.1S細胞に対する様々なスペーサードメインを備えるCD8
+
SLAMF7-CAR T細胞及び形質導入していないT細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=3の独立した実験からの平均±SEMとして表す。C. CD4
+
SLAMF7 CAR T細胞と、K562又はMM.1S標的細胞との24時間の共培養から得られた上清中のIFNγを検出するELISA。T細胞(50,000/ウェル) 及び標的細胞(12,500/ウェル) を、三重測定ウェルに播種した。値はn=2の独立した実験からの平均±SEMとして表す。
IgG3由来スペーサーを備えるROR2 CAR T細胞はin vitroで機能的である。A. 抗原特異的増殖。CD8
+
T細胞をCFSEで標識し、ROR2の発現を伴う又は伴わない照射したMDA-MB231腫瘍細胞を用いて4:1のE:T比で刺激した。CFSEの希釈によって可視化した増殖を、72時間後に判定した。外因性サイトカインは添加しなかった。n=3の独立した実験についての代表的な例。B. 生物発光系アッセイ(3時間のインキュベーション)における、ROR2
+
U266細胞に対する様々なスペーサードメインを備えるCD8
+
ROR2-CAR T細胞及び形質導入していないT細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=3の独立した実験からの平均±SEMとして表す。C. CD8
+
ROR2 CAR T細胞と、ROR2の発現を伴う又は伴わないMDA-MB231標的細胞との24時間の共培養から得られた上清中のIFNγを検出するELISA。T細胞(50,000/ウェル)及び標的細胞(12,500/ウェル)を、三重測定ウェルに播種した。値はn=2の独立した実験からの平均±SEMとして表す。
IgG3由来スペーサーを備えるCD19 CAR T細胞はin vivoで機能的である。NSGマウスに1×10
6
個のRaji細胞(ffluc
+
GFP
+
)を接種し、7日目に5×10
6
個のCD8
+
CD19-CAR T細胞で処置するか又は未処置のままとした。A. 各処置群における白血病の進行/退縮を評価するための連続生物発光イメージング。B. CD19-CAR T細胞(n=3)又は形質導入していないT細胞(n=2)によって処置したマウスの群におけるマウスの生存のKaplan-Meier解析をパネルAに示す。統計解析はログ-ランク検定を用いて行った。* p<0.01。C. 更に6匹のマウスに1×10
6
個のRaji細胞(ffluc
+
GFP
+
)を接種し、7日目に5×10
6
個のCD8
+
CD19-CAR T細胞(IgG3_MiH3バリアント又はIgG4対照CAR)で処置した。7日後にマウスを犠牲死させ、末梢血、骨髄、及び脾臓におけるCAR T細胞の存在について解析した。
IgG3由来スペーサーを備えるROR1 CAR T細胞はin vivoで機能的である。NSGマウスに1×10
6
個のJeko-1細胞(ffluc
+
GFP
+
)を接種し、7日目に5×10
6
個のCD8
+
ROR1-CAR T細胞(R11 scFv)で処置するか又は未処置のままとした。R11 ROR1-CAR T細胞(n=5)又は形質導入していないT細胞(n=5)によって処置したマウスの群における生存のKaplan-Meier解析。
さらなる多機能部位を有するCARの設計及び検出。A. 様々なIgG3スペーサーを備えるCD19 CAR又はIgG4対照CARを形質導入したCD8+ T細胞における抗MiH抗体#1を用いるCAR表面発現の直接染色。B. IgG4参照CAR及び第1世代IgG3スペーサーCAR又はさらなる多機能部位を有する新型IgG3フォーマットの概略説明図。
さらなるMFSを有する新型IgG3 CARフォーマットはin vitro及びin vivoにおいて強力なCD19 CAR T抗腫瘍機能を提供する。A. 抗原特異的増殖。CD8
+
T細胞をCFSEで標識し、CD19の発現を伴う又は伴わない照射したK562腫瘍細胞を用いて4:1のE:T比で刺激した。CFSEの希釈によって可視化した増殖を、72時間後に判定した。外因性サイトカインは添加しなかった。n=3の独立した実験についての代表的な例。B. 生物発光系アッセイ(3時間のインキュベーション)における、CD19
+
Jeko-1細胞に対する様々なスペーサードメイン±scFv V
H
とV
L
との間のさらなるIgG3系多機能部位を備えるCD8
+
CD19-CAR T細胞又は形質導入していないT細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=3の独立した実験からの平均±SEMとして表す。C. CD8
+
CD19 CAR T細胞と、CD19の発現を伴う又は伴わないK562標的細胞との24時間の共培養から得られた上清中のIFNγを検出するELISA。T細胞(50,000/ウェル)及び標的細胞(12,500/ウェル)を、三重測定ウェルに播種した。値はn=2の独立した実験からの平均±SEMとして表す。D. NSGマウスに1×10
6
個のRaji細胞(ffluc
+
GFP
+
)を接種し、7日目に5×10
6
個のCD19-CAR T細胞(CD4
+
:CD8
+
比1:1)で処置するか又は未処置のままとした。各処置群における白血病の進行/退縮を評価するために連続生物発光イメージングを行なった。E. CD19-CAR T細胞(n=5)又は形質導入していないT細胞(n=5)によって処置したマウスの群におけるマウスの生存のKaplan-Meier解析をパネルDに示す。
多機能部位を標的とすることによるCAR T細胞の富化。CD8
+
CAR T細胞を模擬T細胞と1:1の比で混合し、いずれもビオチン化した形態の抗MiH抗体#1又は抗EGFRt抗体で標識した。細胞を洗浄し、抗ビオチン-マイクロビーズで標識し、再び洗浄し、Miltenyi社MACS LSカラムを用いて単離し、翌日にフローサイトメトリーによって解析した。A. IgG3-スペーサーCARS対IgG4フォーマットについてのCD8
+
EGFRt
+
細胞のパーセンテージとしてのソーティング後の陽性分画の純度(n=4の独立した実験)。B. Aにおける富化の効率、陽性分画中のEGFRt
+
細胞の数を陽性+陰性の分画中のEGFRt
+
細胞の全数で除したパーセントとして示す(n=4の独立した実験)。上のパネルは、フローサイトメトリーによって測定したソーティング後のCD19_IgG3_MiH5 CAR T細胞の純度についての例を示す。C. 第1世代IgG3-スペーサーCAR (R11_IgG3_MiH3)又は新型IgG3フォーマット(R11_IgG3_MiH5/MiH3)についてのCD8
+
EGFRt
+
細胞のパーセンテージとしてのソーティング後の陽性分画の純度(n=3の独立した実験)。D. Cにおける富化の効率、陽性分画中のEGFRt
+
細胞の数を陽性+陰性の分画中のEGFRt
+
細胞の全数で除したパーセントとして示す(n=3の独立した実験)。示した値は、培地のみで処理した細胞に対する倍数変化として計算する。
IgG3スペーサーを標的とすることによるCAR T細胞の活性化及び拡大。様々なIgG3系スペーサーを備えるCD8
+
CAR T細胞を、5μg/mlの抗MiH抗体#1でプレコートした96ウェルのプレートに三重測定で播種して24時間(A、B)培養し、続いてCD25(A)及びCD69(B)の発現のフローサイトメトリー解析を行なうか、又は拡大アッセイ(C、上パネル)のために7日間培養し、続いて細胞の計数(C、下パネル)を行なった。アステリスクはWilcoxon検定によって確立された統計的有意を示す。p<0.05。
多機能部位を標的とすることによるCAR Tの増殖の誘起。CD8
+
T細胞をCFSEで標識し、抗IgG3ヒンジCARの発現を伴う若しくは伴わない照射したK562腫瘍細胞(K562_抗CAR)を用いて4:1のE:T比で、又は抗CD3/抗CD28、抗MiH抗体#1、抗MiH抗体#1/抗CD28若しくは抗MiH抗体#1/抗4-1BBでコートしたDynabeads(登録商標)を用いてビーズ対T細胞の比1.6で、刺激した。CFSEの希釈によって可視化した増殖を、72時間後に判定した。外因性サイトカインは添加しなかった。A. 増殖を誘起するために用いた試薬の概略図。B. 第1世代IgG3-スペーサーCAR(CD19_IgG3_MiH1(左パネル))、又は新型IgG3フォーマット(CD19_IgG3_MiH5/MiH1、右パネル)についてのn=3の独立した実験からの代表的な例。
IgG3スペーサーを標的とすることによるCAR T細胞のADC媒介枯渇。様々な長さのIgG3由来スペーサー又はIgG4参照CARを備える5×10
4
個のCD8
+
CAR T細胞、並びに形質導入していないT細胞を三重測定ウェルに播種し、様々な濃度の抗MiH抗体#1-ADC(アンスラサイクリン系細胞傷害性ペイロードにコンジュゲートした抗MiH抗体#1)で処理した。50IUのIL-2の存在下に細胞を72時間培養し、洗浄して、フローサイトメトリー解析に供した。示した生存細胞のパーセンテージは、処理した試料の正規化した細胞数を、培地のみで培養したそれぞれの細胞の正規化した細胞数で除したものである(n=5の独立した実験)。
多機能部位をin vitroで標的とすることによるCAR T細胞のADC媒介枯渇。最適化されたIgG3スペーサーバージョン、最適化されたIgG3スペーサーバージョン+ scFv V
H
とV
L
との間のさらなる多機能部位、又はIgG4参照CARを備える5×10
4
個のCD8
+
CAR T細胞、並びに形質導入していないT細胞を三重測定ウェルに播種し、様々な濃度の抗MiH抗体#1-ADC(アンスラサイクリン系細胞傷害性ペイロードにコンジュゲートした抗MiH抗体#1)で処理した。50IUのIL-2の存在下に細胞を72時間培養し、洗浄して、フローサイトメトリー解析に供した。示した生存細胞のパーセンテージは、処理した試料の正規化した細胞数を、培地のみで培養したそれぞれの細胞の正規化した細胞数で除したものである。A. CD19 CARを用いるADCアッセイ(CD19_IgG3_MiH1対CD19_IgG3_MiH5/MiH1対CD19_IgG4)。n=3の独立した実験からのデータ。B. CD20 CARを用いるADCアッセイ(CD20_IgG3_MiH3対CD20_IgG3_MiH5/MiH3対CD20_IgG4)。n=2の独立した実験からのデータ。C. ROR1 CARを用いるADCアッセイ(R11_IgG3_MiH3対R11_IgG3_MiH5/MiH3対R11_IgG4)。n=2の独立した実験からのデータ。
抗IgG3ヒンジCARでin vitroで標的とすることによるCAR発現細胞の枯渇。生物発光系アッセイ(5時間のインキュベーション)における、CD19-CAR CD19_MiH5を形質導入したK562細胞(A)又はK562細胞(B)に対する抗CAR(IgG4スペーサーを有する抗MiH抗体#1系CAR)を備えるCD8
+
T細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=3の独立した実験からの平均±SEMとして表す。
in vivoにおけるCAR T細胞のADC媒介枯渇。NSGマウスに、新型バージョンのIgG3系CD19 CAR(CD19_IgG3_MiH5/MiH1)並びにffluc_GFP融合タンパク質を形質導入した4.5×10
6
個のCD4
+
T細胞を接種した。8日目にマウスの半分(1群あたり動物n=8)を100μgの抗MiH抗体#1 ADC(4.5mg/kg体重に対応)で処置した。11日目に、抗MiH抗体#1系抗CARを備える照射したK562細胞(マウスあたり1×10
6
の照射したK562_抗CAR細胞)でT細胞を再刺激した。A. 連続生物発光イメージングによってT細胞の持続性の速度論を評価した。B. 18日目におけるエンドポイント生物発光。
in vivoにおけるCAR T細胞のCAR特異的刺激。NSGマウスに、増殖染料eFluor670で標識した新型バージョンのIgG3系CD19 CAR(CD19_IgG3_MiH5/MiH1)を形質導入した4.5×10
6
個のCD4
+
T細胞を接種した。T細胞を移送した後、続いてn=5のマウスの群を様々な時点で3×10
6
個の照射したK562又はK562_抗CAR細胞で処置した。1つの群にはT細胞の注射の日(d0)のT移送の3時間後にK562_抗CAR細胞を投与した。第2の群にはT細胞の注射の後3日目に、照射したK562_抗CAR細胞のさらなる投薬を行なった(d0+d3)。他の2群はそれぞれ、T細胞の移送後1日目(d1)又はd1+d3に、照射したK562_抗CAR細胞で処置した。対照群には照射したK562細胞をd0+d3に投与した。T細胞の移送後4日目にマウスを犠牲死させ、骨髄細胞を収集した。CD4、CD45、及びEGFRtに対する抗体で細胞を染色し、フローサイトメトリー解析に供した。CD45
+
/CD4
+
/EGFR
+
骨髄由来T細胞を、eFluor 670希釈について解析した。
さらなるMFSを有する新型IgG3 CARフォーマットはin vitroで強力なROR1 CAR T抗腫瘍機能を提供する。A. 抗原特異的増殖。CD8
+
T細胞をCFSEで標識し、ROR1の発現を伴う又は伴わない照射したK562腫瘍細胞を用いて4:1のE:T比で刺激した。CFSEの希釈によって可視化した増殖を72時間後に判定した。外因性サイトカインは添加しなかった。n=3の独立した実験についての代表的な例。B. 生物発光系アッセイ(5時間のインキュベーション)における、ROR1
+
K562_ROR1細胞に対する様々なスペーサードメイン±scFv V
H
とV
L
との間のさらなるIgG3系多機能部位を備えるCD8
+
ROR1-CAR T細胞又は形質導入していないT細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=4の独立した実験からの平均±SEMとして表す。C. CD8
+
ROR1 CAR T細胞と、ROR1の発現を伴う又は伴わないK562標的細胞との24時間の共培養から得られた上清中のIFNγを検出するELISA。T細胞(50,000/ウェル)及び標的細胞(12,500/ウェル)を、三重測定ウェルに播種した。値はn=3の独立した実験からの平均±SEMとして表す。
さらなるMFSを有する新型IgG3 CARフォーマットはin vitroで強力なCD19 CAR T抗腫瘍機能を提供する。A. 抗原特異的増殖。CD8
+
T細胞をCFSEで標識し、CD19の発現を伴う又は伴わない照射したK562腫瘍細胞を用いて4:1のE:T比で刺激した。CFSEの希釈によって可視化した増殖を72時間後に判定した。外因性サイトカインは添加しなかった。n=3の独立した実験についての代表的な例。B. 生物発光系アッセイ(3時間のインキュベーション) における、CD19
+
Jeko-1細胞に対する様々なスペーサードメイン±scFv V
H
とV
L
との間のさらなるIgG3系多機能部位を備えるCD8
+
CD19-CAR T細胞又は形質導入していないT細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=3の独立した実験からの平均±SEMとして表す。C. CD8
+
CD19 CAR T細胞と、CD19の発現を伴う又は伴わないK562標的細胞との24時間の共培養から得られた上清中のIFNγを検出するELISA。T細胞(50,000/ウェル)及び標的細胞(12,500/ウェル)を、三重測定ウェルに播種した。値はn=3の独立した実験からの平均±SEMとして表す。
さらなるMFSを有する新型IgG3 CARフォーマットはin vitroで強力な細胞傷害性効果を提供する。生物発光系アッセイにおける、抗原
+
腫瘍細胞に対する様々なスペーサードメイン±scFv V
H
とV
L
との間のさらなるIgG3系多機能部位を備えるCD8
+
CAR T細胞又は形質導入していないT細胞の特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値はn=2の独立した実験からの平均±SEMとして表す。A. ROR1
+
Jeko-1細胞に対するROR1特異的CAR T細胞(4-2 scFv) (3時間のインキュベーション)。B. FLT3
+
MOLM-13細胞に対するFLT3特異的CAR T細胞(4G8 scFv) (5時間のインキュベーション)。C. FLT3
+
MOLM-13細胞に対するFLT3特異的CAR T細胞(BV10 scFv) (5時間のインキュベーション)。D. Siglec-6
+
MV4-11細胞に対するSiglec-6特異的CAR T細胞(JML-1 scFv) (3時間のインキュベーション)。
さらなるMFSを有する新型IgG3 CARフォーマットはin vitroで強力なCD19 CAR T抗腫瘍機能を提供する。A. NSGマウスに1×10
6
個のRaji細胞(ffluc
+
GFP
+
)を接種し、7日目に5×10
6
個のCD19-CAR T細胞(CD4+:CD8+比1:1)で処置するか又は未処置のままとした。各処置群における白血病の進行/退縮を評価するために連続生物発光イメージングを行なった。B. NSGマウスに1×10
6
個のRaji細胞(ffluc
+
GFP
+
)を接種し、7日目に5×10
6
個のCD19-CAR T細胞(CD4+:CD8+比1:1)で処置するか又は未処置のままとした。各処置群における白血病の進行/退縮を評価するために連続生物発光イメージングを行なった。C. D19-CAR T細胞(n=7)又は形質導入していないT細胞(n=4)によって処置したマウスの群におけるマウスの生存のKaplan-Meier解析をパネルA~Bに示す。
IgG3 MFSを標的とすることによるin vitroにおけるCAR T細胞の拡大。5×10
5
個のCD4
+
若しくはCD8
+
の形質導入していない対照T細胞、又は新型IgG3フォーマットのCD19特異的CARを備えるCD4
+
若しくはCD8
+
のT細胞を、5×10
6
個の照射したTM-EBV-LCL(CD19
+
)若しくはK562_抗CAR(CD19
-
)フィーダー細胞及び50IUのIL-2の存在下で14日間拡大させ、14日間の拡大の後にT細胞を計数した。形質導入していないT細胞のみについては、拡大の設定に30ng/mlのOKT3が含まれていた。値は、開始時の計数に対して14日後のx倍拡大として表す(n=2の独立した実験からの平均±SEM)。
IgG3 MFSを標的とすることによるin vivoにおけるCAR T細胞の拡大。NSGマウスにCD19-CAR CD19_MiH5/MiH1を形質導入した1×10
7
個のffluc
+
GFP
+
T細胞を接種し(CD4
+
:CD8
+
比2.7:1)、8日目に1×10
7
個のK562_抗CAR細胞又は形質導入していない対照のK562細胞で処置した。処置群におけるT細胞の持続性/拡大を評価するため、連続生物発光イメージングを行なった。
in vitro及びin vivoにおいて抗IgG3ヒンジCARで標的とすることによるCAR発現細胞の枯渇。A~D. 生物発光系アッセイ(5時間のインキュベーション)における、抗CAR(IgG4スペーサーを有する抗MiH抗体#1系CAR)を備える3匹の異なるドナーからのCD8
+
T細胞の、ホタルルシフェラーゼ(A、C)又はホタルルシフェラーゼ及び抗CD19-CAR CD19_MiH5/MiH1(B、D)を形質導入した同じ3匹のドナーからのCD4
+
T細胞に対する特異的細胞溶解活性。アッセイは、三重測定ウェルで標的細胞5,000個/ウェルとして実施した。値は、それぞれドナー1(A、B)及びドナー2(C、D)からのCD8
+
エフェクター抗CAR T細胞を用いたn=1の実験からの三重測定ウェルの平均として表す。E~F. 1群あたり4匹のNSGマウスにドナー2からの2.2×10
6
個の標的T細胞(CD4
+
:CD8
+
比1:1) (ffluc
+
GFP
+
+ 抗CD19-CAR CD19_MiH5/MiH1)を接種し、24時間後に4×10
6
個のCD8
+
抗CAR-CAR T細胞(ドナー2)又は同じドナーからの形質導入していない対照T細胞で処置した。各処置群におけるT細胞の持続性/枯渇を評価するため、連続生物発光イメージングを行なった。注:1日目のみの形質導入していない群の左側のマウスは、実験に含めなかった。
【発明を実施するための形態】
【0008】
過去数十年の間に、キメラ抗原受容体(CAR) (以前はT-Bodyとも呼ばれた)の設計は、やや単純な構築物から、異なるタンパク質ドメインから組み立てられるより複雑な分子へと進化した。今日では第一世代CARとも呼ばれるその最も単純な形態では、CARはCD3ζサブユニットのシグナル伝達ドメインに融合されたモノクローナル抗体のscFvからなった
1
。その後、完全なT細胞エフェクター機能を誘導するには、ほとんどのCAR構築物がscFvと膜貫通ドメインの間にスペーサーを必要とすることが明らかになった
2
。1990年代半ばから現在まで、Fc領域又は免疫グロブリン様ドメインは異なるタンパク質(CD4、CD7、CD8α、CD28、IgD、IgG1及びIgG4を含み、CD8αヒンジが最も一般的に使用され、最もよく調べられているものである
3-6
)に由来したが、当分野のほとんどの研究者はそのCAR構築物の全てにスペーサー1つのみのフォーマットを使用している。使用されるCAR設計は、最も機能的なものとは限らない場合があるにもかかわらず、これは有効な結果をもたらした。異なる抗体は、その標的分子上の異なるエピトープを結合する。本発明者らは以前の研究において、標的エピトープに最適に適合するように組成及び長さを調整されたスペーサーが最大の抗腫瘍機能をもたらすことを示した
7
。
【0009】
本発明は、遺伝子改変免疫受容体に組み込む、例えば、CAR構築物にスペーサードメインとして組み込むための、ヒトIgG3のヒンジドメインの新規バリアントを提供する。
【0010】
本発明者らは、scFv及び膜貫通ドメインがヒトIgG3ヒンジドメインのバリアントによって連結された、IgG3由来スペーサーを有するCARのライブラリーを生成した。これは、天然には上部ヒンジ(12aa、ELKTPLGDTTHT、配列番号2)、中央部ヒンジ(50aa、CPRCP、配列番号59 + 15aaモチーフEPKSCDTPPPCPRCP、配列番号1の3リピート)及び下部ヒンジ(8aa、APELLGGP、配列番号60)からなり、IgG3_UMLH (上部、中央部及び下部ヒンジ)と呼ばれるこの野生型スペーサーに70aaの総スペーサーサイズをもたらす。本発明者らは、上部ヒンジ(ELKTPLGDTTHT、配列番号2)からなるバリアントを構築したことから、中央部ヒンジのn末端部分(CPRCP、配列番号59)及びEPKSCDTPPPCPRCPモチーフ(配列番号1)の0~10コピーは、IgG3_MiH0~IgG3_MiH10と称する、15aaステップ中17~167aaに及ぶスペーサードメインをもたらした。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
株式会社トクヤマ
四塩化炭素の製造方法
1か月前
東ソー株式会社
炭素-窒素結合形成方法
1か月前
株式会社トクヤマ
シロキサン類の回収方法
1か月前
日産化学株式会社
ピリジン化合物の製造方法
3か月前
株式会社トクヤマ
ビオチン誘導体の製造方法
20日前
株式会社コスモス
液状炭化水素の増産方法
4日前
株式会社トクヤマ
ベンザゼピン化合物の製造方法
3日前
株式会社コスモス
液状炭化水素の増産方法
5日前
日本特殊陶業株式会社
メタン製造装置
2か月前
ユニチカ株式会社
ビスマレイミドおよびその製造方法
2か月前
四国化成工業株式会社
エポキシ化合物およびその利用
2か月前
信越化学工業株式会社
新規化合物
1か月前
artience株式会社
四塩基酸無水物の製造方法
1か月前
金剛化学株式会社
ボルチオキセチンの製造方法
2か月前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
18日前
日産化学株式会社
ピラゾール化合物及び有害生物防除剤
1か月前
四国化成工業株式会社
テレフタル酸化合物およびその利用
2か月前
株式会社トクヤマ
サフィナミド若しくはその塩の製造方法
1か月前
四国化成工業株式会社
イソフタル酸化合物およびその利用
2か月前
株式会社カネカ
プロピレンオキサイド(PO)製造システム
2か月前
旭化成株式会社
トリオキサンの製造方法
1か月前
キヤノン株式会社
有機化合物及び有機発光素子
1か月前
東ソー株式会社
免疫グロブリン結合性タンパク質の保存溶液
1か月前
JNC株式会社
有機ケイ素化合物およびこれを用いた重合体
4日前
キヤノン株式会社
有機化合物及び有機発光素子
2か月前
キヤノン株式会社
有機化合物及び有機発光素子
2か月前
キヤノン株式会社
有機化合物及び有機発光素子
2か月前
株式会社半導体エネルギー研究所
有機金属錯体、発光デバイス
2か月前
JFEスチール株式会社
メタノール合成方法
2か月前
旭化成株式会社
ホルムアルデヒドの製造方法
2か月前
公立大学法人大阪
カルボン酸化合物の製造方法
1か月前
株式会社トクヤマ
精製第4級アンモニウム化合物水溶液の製造方法
3か月前
株式会社半導体エネルギー研究所
有機化合物、および発光デバイス
1か月前
住友ベークライト株式会社
フェノールの製造方法
2か月前
学校法人東海大学
樹脂担持環状テルル化合物
18日前
東京都公立大学法人
キチン系複合体
11日前
続きを見る
他の特許を見る







 特許ウォッチ
特許ウォッチ