TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2024054422
公報種別
公開特許公報(A)
公開日
2024-04-16
出願番号
2024030532,2021106286
出願日
2024-02-29,2021-06-28
発明の名称
感染性植物ラブドウイルスベクターおよび植物における非トランスジェニックのゲノム部位特異的編集の方法
出願人
浙江大学
,
ZHEJIANG UNIVERSITY
代理人
個人
主分類
C12N
15/09 20060101AFI20240409BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】植物ラブドウイルスベクターを用いて植物染色体DNAを部位特異的修飾する方法およびウイルスベクターの組成物を提供する。
【解決手段】植物ラブドウイルスベクターを用いて植物を全身感染し、配列特異的ヌクレアーゼを発現し、配列特異的ヌクレアーゼが、感染された植物ゲノム標的サイトを標的とし、標的サイトを切断するステップと、植物のDNA修復メカニズムにより標的サイトに対する部位特異的編集を完了するステップを含む。この植物ゲノム目的DNAを部位特異的編集する方法は、選択的マーカー遺伝子を導入する必要がない。植物ラブドウイルス発現ベクターを用いて配列特異的ヌクレアーゼを送達することにより、標的サイトに対する編集効率を向上させるだけでなく、非トランスジェニックの安定に遺伝可能な編集植株を獲得することができる。
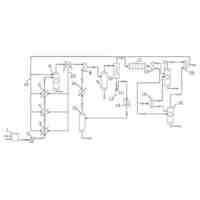
【選択図】図1
特許請求の範囲
【請求項1】
修飾すべき細胞ゲノム内に外来遺伝子配列を導入する必要がない植物細胞遺伝物質を修飾する方法であって、
少なくとも1つの遺伝物質修飾すべき植物細胞を提供するステップa)と、全身感染能力を有する組み換え植物ラブドウイルスベクターを用いて前記植物細胞を感染するステップであって、前記全身感染能力を有する組換え型ラブドウイルスは、少なくとも1つの配列特異的エンドヌクレアーゼをコードするリボ核酸配列を担持し、前記配列特異的ヌクレアーゼは、植物ゲノム核酸配列を特異的に標的とし、標的サイトを切断するステップb)とを含み、
前記細胞遺伝物質修飾プロセスは、配列特異的ヌクレアーゼによって切断誘導され、植物内因性DNA修復機器によって完成される、植物細胞遺伝物質を修飾する方法。
続きを表示(約 1,300 文字)
【請求項2】
修飾すべき植物細胞ゲノム内に外来遺伝子配列を導入する必要がない遺伝物質が修飾された植物を産生する方法であって、
少なくとも1つの遺伝物質修飾すべき植物細胞を提供するステップa)と、全身感染能力を有する組み換え植物ラブドウイルスベクターを用いて前記植物細胞を感染するステップであって、前記全身感染能力を有する組換え型ラブドウイルスは、少なくとも1つの配列特異的エンドヌクレアーゼをコードするリボ核酸配列を担持し、前記配列特異的ヌクレアーゼは、植物ゲノム核酸配列を特異的に標的とし、標的サイトを切断するステップb)と、前記遺伝物質が修飾された細胞から植物を取得するステップc)、選択マーカーを利用する必要がなく、前記遺伝物質を含む修飾植物対象を選択するステップd)とを含む、遺伝物質が修飾された植物を産生する方法。
【請求項3】
前記修飾すべき細胞を感染する方法は、組み換えウイルス未接種細胞自然感染、汁の摩擦接種、接ぎ木、昆虫媒介伝播、または他の任意のウイルスによる感染方法を含む、ことを特徴とする請求項1に記載の方法。
【請求項4】
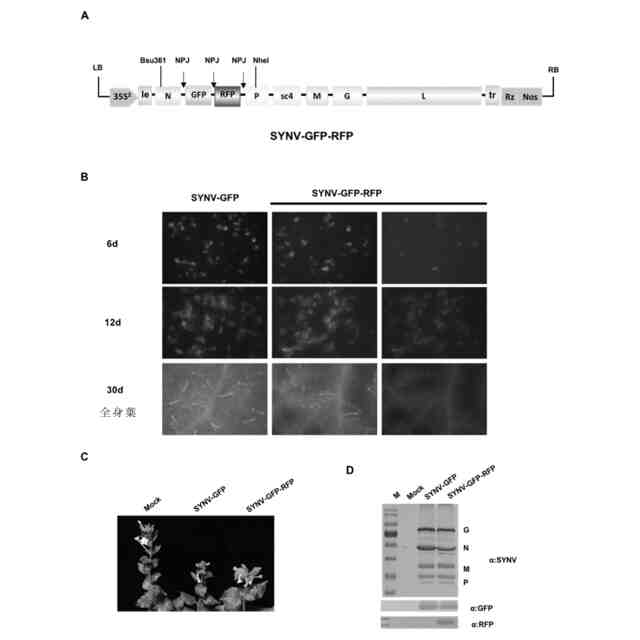
前記全身感染能力を有するラブドウイルスベクターが、ノゲシ黄網ウイルス(sonchus yellow net virus、SYNV)またはナス斑点萎縮ウイルス(eggplant mottled dwarf virus、EMDV)である、ことを特徴とする請求項1に記載の方法。
【請求項5】
前記全身感染能力を有するラブドウイルスベクターは、一つまたは複数の種類の突然変異および/または組み換え修飾後の誘導ウイルスベクターを含む、ことを特徴とする請求項1に記載の方法。
【請求項6】
前記誘導ウイルスベクターが、糖タンパク質(Glycoprotein、G)アミノ基末端ドメインが突然変異された、および/または組み換え修飾後の誘導ウイルスベクターである、ことを特徴とする請求項5に記載の方法。
【請求項7】
前記細胞遺伝物質修飾は、標的サイトで少なくとも1つの塩基欠失、または1つの塩基挿入、または1つの塩基置換、またはこれらの修飾パターンの組み合わせから選択される、ことを特徴とする請求項1に記載の方法。
【請求項8】
前記配列特異的エンドヌクレアーゼは、CRISPR/Casヌクレアーゼ、TALENヌクレアーゼ、ジンクフィンガーヌクレアーゼ、またはすべてのゲノム編集を実現可能なヌクレアーゼである、ことを特徴とする請求項1に記載の方法。
【請求項9】
前記配列特異的エンドヌクレアーゼをコードするリボ核酸配列は、単一または複数のガイドRNAの核酸配列と、Casヌクレアーゼの核酸配列とを含む、ことを特徴とする請求項1に記載の方法。
【請求項10】
前記配列特異的ヌクレアーゼ配列は、ラブドウイルスのゲノム中に独立した転写ユニットの形態で存在する、ことを特徴とする請求項9に記載の方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明は、植物遺伝子工学技術分野に関し、特に、組換え型ラブドウイルスベクターを用いて植物に対して部位特異的編集を行う方法に関し、具体的には、本発明は、組換え型ラブドウイルスベクターを用いてCRISPRヌクレアーゼ成分を非トランスジェニック送達することで植物に対して部位特異的編集を行う方法に関する。
続きを表示(約 4,500 文字)
【背景技術】
【0002】
近年、ゲノム部位特異的編集技術(Targeted Genome Editing)の出現は、作物の精密育種のために新規ルートを開拓し、小麦、水稲、トウモロコシなどの多種の作物の機能ゲノム学研究および性状の遺伝改良において非常に広い応用展望を有する。現在、ゲノム部位特異的編集技術は、主に配列特異的ヌクレアーゼ(site-specific nucleases, SSNs)に依存して完成している。配列特異的ヌクレアーゼとは、部位特異的なDNA結合ドメイン(DNA binging domain)と非特異的のDNA切断ドメイン(DNA cleavage domain)とを組み合わせ、ゲノム特定領域を標的かつ改造可能なドメインである。配列特異的ヌクレアーゼを細胞に導入された後に、ゲノムにおける特異配列を識別し、当該配列を切断してDNAの二重鎖切断(double-stranded breaks、DSBs)を形成することができ、それによって細胞内の非相同末端結合(non-homologous end joining、NHEJ)または相同組換え(homologous recombination、HR)を活性化して修復し、遺伝子機能の欠失(Deletion)或いは置換(Replacement)を引き起こす。現在よく使われる配列特異的ヌクレアーゼは、主にジンクフィンガーヌクレアーゼ(zinc-finger nucleases、ZFN)、転写活性化因子様エフェクターヌクレアーゼ(transcription activator-like effector nucleases、TALEN)、クラスター化して規則的な短い配列の回文配列リピートおよびCRISPR関連システム(clustered regularly interspaced short palindromic repeats/CRISPR associated proteins、CRISPR/Cas)という3種類を含む。そのうちのCRISPR/Casヌクレアーゼは、操作が簡便で、迅速かつ効率が高い利点があり、ゲノム編集ツールとして現在最も広く応用されている。
【0003】
現在の植物ゲノム編集のキーテクノロジーの1つは、配列特異的ヌクレアーゼの植物送達技術であり、すなわち、配列特異的ヌクレアーゼを含む核酸分子および/又はタンパク質分子を標的細胞、組織、器官又は植株個体に導入する方法と手段である。理想的な送達技術として、以下の条件を満たさなければならない:(1)完全な配列特異的ヌクレアーゼ核酸やタンパク質分子を植物材料に導入することができる;(2)送達される受容体植物材料が獲得しやすく、導入過程が簡単で操作しやすい;(3)植物材料中に導入した配列特異的ヌクレアーゼは、高効率の編集効率を発揮できる;(4)編集後の突然変異再生植株が獲得しやすい。(5)獲得した編集植株は、外来核酸の統合を含まない。
【0004】
従来技術では、CRIPSR/Casヌクレアーゼを植物に送達することは、主に遺伝子組換え方法、すなわち、CRIPSR/Casを含む組換え遺伝子を受容体植物細胞に導入して植物のゲノムに統合し、発現産生されたヌクレアーゼがゲノム標的配列に対して予想可能な指向性遺伝的変化を産生することに依存する。現在、植物において最もよく使われている技術は、アグロバクテリウム介在法と遺伝子銃法などを含み、ゲノムに関する外来DNA挿入ということはすべて遺伝子組換えとして見なされる。当該方法は、ヌクレアーゼの植物への送達を実現することができるが、依然として、育種応用において以下の問題が存在する:(1)植物ゲノムにおける外来遺伝子の統合に関し、また、形質転換の過程では抗生物質や除草剤を用いてマーカーをスクリーニングする必要がある;(2)獲得した編集植株に予期できない表現型変異が存在する可能性がある。(3)自配または戻し交配などの手段により組換え遺伝子を分離する過程周期が長く、特に、育種周期が長く、倍数体、および無性生殖の植物である場合;(4)ある国や地区では、組換え遺伝子を除去した編集植株の子の世代およびその製品は、遺伝子組換え製品の監督管理対象となる可能性がある。
【0005】
遺伝子組換え送達方式に問題が存在する状況に鑑み、非トランスジェニックCRIPSR/Casヌクレアーゼの送達方式を構築する必要がある。植物ウイルスベクターは、もう一つの将来性のあるCRISPR/Casヌクレアーゼ送達方法である。全身感染能力を有するウイルスベクターは、接種された細胞または組織から他の未接種組織および細胞に拡張することができ、かつ植株個体レベルで外来遺伝子の全身送達を達成することができる。全身感染能力を有さないウイルスベクターは、初感染細胞から他の細胞に拡散することができず、したがって、全身送達を達成することができない。全身感染能力を有するウイルスベクター、特に感染性RNAウイルスベクターは、その複製過程がDNA段階に関連しないため、外来核酸が寄主ゲノムに統合される可能性が存在しなく、ヌクレアーゼの非トランスジェニック送達応用において特殊な価値を有する。しかしながら、CRISPR/Casヌクレアーゼ遺伝子分子量が比較的大きく、4.2千塩基対(Kb)に達するため、プロモーターエレメントを加えて5-7 kbに達することが可能性があり、現在のウイルスベクターのパッケージング能力に対して極めて大きな挑戦を提出した。外来遺伝子の長さが感染性ウイルスベクターの限界負荷能力を超えた場合、外来遺伝子フラグメントの紛失、欠失をもたらすか、またはウイルスの全身感染能力を喪失する(Avesaniら、Transgenic Res、2007、16:587-597)。
【0006】
パッケージング能力の制約のため、一部の感染性植物ウイルスベクターは、長さ約100~200塩基対(bp)のCRISPRガイドRNA(gRNA)などの比較的小さいヌクレアーゼ成分を送達するためにのみ使用され、このようなベクターは、例えば、植物DNAウイルス中のキャベツ葉巻ウイルス(Yinら、Sci Rep、2015、5:14926)、プラスセンスRNAウイルス中のタバコ茎壊疽ウイルス(Aliら、Mol Plant、2015、8:1288-1291;WO 2015189693)、タバコモザイクウイルス(Codyら、Plant Physiol、2017、175:23-35)、エンドウ豆早期褐変ウイルス(Aliら、Virus Res、2018、244:333-337)、テンサイそう根病(Jiangら、Plant Biotechnol J、2019、17:1302-1315)などがある。これらのウイルスベクターは、Cas9遺伝子組換え植株を介してゲノム部位特異的編集を実現するのに必要とされるため、このようにして獲得した編集植株のゲノムにCas9核酸配列の挿入がまだ含まれている。
【0007】
論文(Meiら、Plant Direct、2019、3:1-16)では、約700bpの緑色蛍光タンパク質遺伝子を持つ場合、感染した上部葉で外来挿入フラグメントが完全欠失し、100~200bpのCRISPR gRNAを発現するためにのみ利用可能となり、遺伝子組換え発現のCas9タンパク質に依存して遺伝子編集を実現可能なプラスセンスRNAウイルスに属するオヒゲシバモザイク ウイルスウイルスベクターが開示される。論文(Zhangら、Funct&Integr Genomics、2020、20:471-477)および中国特許CN 111979262 Aと同様に、当該ウイルスベクターを用いて植物ゲノム編集を行う方法が開示される。パッケージング能力の制約のため、当該がCas9とgRNAをそれぞれ2つのウイルスベクターに挿入し、植物を共同に感染するときに低頻度の遺伝子編集を実現した。
【0008】
もう1つのプラスセンスRNAウイルスに属するジャガイモXウイルスの外来フラグメントに対する許容能力は、2 kb未満であった(Larsenら、BMC Biotech、2012、12:21;Avesaniら、Transgenic Res、2007、16:587-597)。論文(Arigaら、Plant Cell Physiol、2020、61:1946-1953)では、当該ベクターを用いてCRISPR/Casヌクレアーゼ遺伝子を送達する方法が開示される。当該ウイルスベクターは、接種葉で完全なCRISPR/Casヌクレアーゼを発現し、遺伝子編集を実現することができるが、未接種組織でCRISPR/Cas全身送達と遺伝子編集を実現することができない。
【0009】
ウイルスレプリコン(Replicon)システムは、ウイルスを複製するために必要とされるエレメントを保留しているが、ウイルスパッケージおよび/または運動に関連するエレメントを除去したベクターである。当該種類のベクターは、効率的に複製できるが、全身感染することができないため、かなり大きい(>10 kb)の外来挿入フラグメントを収容することができる。論文(Baltesら、Plant Cell、2014、151-163)では、一本鎖DNAジェミニウイルス科に属するダイズわい化ウイルスレプリコンを用いてCRISPR/Casヌクレアーゼを送達することが開示されており、論文(Wangら、Mol Plant、2017、10:1007-1010)および中国特許CN 109880846 Aは、ジェミニウイルス科のコムギ萎縮ウイルスレプリコンベクターシステムを用いてCRISPR/CasヌクレアーゼおよびDNA修復テンプレートを送達することが開示される。しかし、ウイルスレプリコンベクターが、実質的に運動欠陥ウイルス(Deconstructed Virus)であり、アグロバクテリウム介在または遺伝子銃などの手法を利用してウイルスベクターを含むDNAコンストラクトを細胞に導入する必要があるので、受容体細胞に外来DNA統合可能性の存在は避けられない。
【0010】
現在すでに開発された植物ウイルスベクターに欠陥が存在する課題に鑑み、感染性を有し、完全なCRISPR/Casヌクレアーゼを全身に送達できる植物ウイルスベクターシステム、特に、複製過程でDNA段階に関しない感染性RNAウイルスベクターシステムを開発する必要がある。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
浙江大学
水平軸方向角振動装置
4か月前
浙江大学
可変粘性流れ抵抗構造
1か月前
浙江大学
超狭線幅レーザ発生装置
1か月前
浙江大学
腹腔内血管自動結紮装置
8か月前
浙江大学
ベース歪み発生装置およびベース歪み感度計測システム
2か月前
浙江大学
高鉄含有量の非晶質ナノ結晶軟磁性合金を調製するための過冷却凝固法
5か月前
浙江大学
複数の変数を同期的にモニタリングする地滑りモデル試験方法及び装置
1か月前
浙江大学
個人化の下肢リハビリテーション歩行パターンの生成装置及び制御方法
4か月前
浙江大学
下肢リハビリテーション訓練用の外骨格の歩行パラメータの最適化方法
4か月前
個人
多軸3Dプリントに基づく自己支持構造の最適化設計及び製造方法
10か月前
浙江大学海南研究院
水稲のカーボンフットプリント評価方法、システム、電子機器及び記憶媒体
9か月前
浙江大学
4Dプリントに基づく迷路状インターロックスマート構造及びそのロック方法
1か月前
個人
構造レイアウト、幾何学的及び3Dプリントの一体型最適化設計及び製造方法
10か月前
海南浙江大学研究院
プロバイオティクスコンビネーションを豚回腸炎予防用生物学的薬剤の製造に使用する方法
9か月前
浙江大学嘉興研究院
メタノール改質と高効率な膜分離との連結に基づくコンパクト型船舶用炭素回収のシステム、方法及び応用
1か月前
浙江大学
感染性植物ラブドウイルスベクターおよび植物における非トランスジェニックのゲノム部位特異的編集の方法
10か月前
湖北万潤新能源科技股フン有限公司
ナトリウム電池用正極材料及びその製造方法
5か月前
他の特許を見る






 特許ウォッチ
特許ウォッチ