TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025094131
公報種別
公開特許公報(A)
公開日
2025-06-24
出願番号
2025045517,2022180974
出願日
2025-03-19,2015-03-19
発明の名称
多能性細胞に関連する方法
出願人
ブイセル セラピューティックス,インコーポレイテッド
代理人
個人
,
個人
,
個人
,
個人
,
個人
主分類
C12N
5/071 20100101AFI20250617BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】効率、収率および/または品質が改善された多能性細胞の生成方法を提供する。
【解決手段】本発明は、細胞からの多能性細胞の作製または生成に関する。本発明は、ストレスが、細胞に外来遺伝子も、転写産物も、タンパク質も、核成分も、細胞質も導入することを必要とせず、また細胞融合も必要とせずに細胞からの多能性幹細胞の産生を誘導し得るという本発明者らの発見に基づくものである。いくつかの実施形態では、ストレスが細胞の細胞質および/またはミトコンドリアの量の減少を誘導し、脱分化過程を惹起して多能性細胞を生じさせる。
【選択図】図5A
特許請求の範囲
【請求項1】
図面に記載の発明。
発明の詳細な説明
【技術分野】
【0001】
(関連出願の相互参照)
本願は、米国特許法第119条(e)の下、2014年3月19日に出願された米国仮特許出願第61/955,362号、2014年3月19日に出願された同第61/955,358号および2014年8月28日に出願された同第62/043,042号の利益を主張するものであり、上記仮特許出願はその全体が参照により本明細書に組み込まれる。
続きを表示(約 11,000 文字)
【0002】
本明細書に記載される技術は多能性細胞の作製に関するものである。
【背景技術】
【0003】
多能性細胞を得るために現在用いられている方法は主として、入手可能性が限られている組織(例えば、胚組織または臍帯血)または外来核酸を導入するリプログラミング因子の添加に依存するものである(Hanna,J.ら,Cell 2008 133,250-264;Hockemeyer,D.ら,Cell stem cell 2008
3,346-353;Kim,D.ら,Cell stem cell 2009 4,472-476;Kim,J.B.Nature 2009 461,649-643;Okabe,M.ら.Blood 2009 114,1764-1767)。外来リプログラミング因子の添加による合併症を起こさない幹細胞、特に自家幹細胞を容易に作製する方法があれば、細胞分化の研究および幹細胞ベースの治療法の開発が加速されるものと思われる。熱傷、化学傷害、外傷および放射線照射などの刺激への曝露により細胞が損傷を受けると、正常な体細胞が変化して癌細胞になる可能性があるという仮説が提唱されているが、リプログラミング因子を特異的に操作しなくても健常成体の体細胞が他の状態に変換され得ることを直接示す証拠はない。
【0004】
これまでに、研究者は成体組織に「成体幹細胞」を発見したことを報告している(Reynolds,B.A.およびWeiss,S.Science 1992 255,1707-1710;Megeney,L.A.ら,Genes & development 1996 10,1173-1183;Caplan,A.I.Journal of orthopaedic research 1991 9,641-650;Lavker,R.M.およびSun,T.T.The Journal of investigative dermatology 1983 81,121s-127s)。このような報告には未だ議論の余地がある。例えば、幹細胞マーカーであるOct4を発現する細胞を探求している研究者で、ホメオスタシスが正常な成体骨髄にOct4発現細胞を発見するには至っていない研究者がいる(Lengner,C.J.ら,Cell
Cycle 2008 7,725-728;Berg,J.S.およびGoodell,M.A.Cel stem cell 2007 1,359-360)一方、様々な成体組織からOct4発現細胞を単離することが可能であることを報告している研究者もいる(Jiang,Y.ら,Nature 2010 418,41-49;D’Ippolito,G.ら,Journal of cell science 2004 117,2971-2981;Johnson,J.ら,Cell 2005 122,303-315;Kucia,M.ら,Leukemia 2006 20,857-869;Kuroda,Y.ら,PNAS 2011 107,8639-8643;Obokata,H.ら,Tissue engineering.2011 Part A
17,607-615;Rahnemai-Azar,A.ら,Cytotherapy 2011 13,179-192;Huang,Y.ら,Transplantation 2010 89,677-685;Zuba-Surma,E.K.ら,Journal of cellular and molecular medicine2011 15,1319-1328;Paczkowska,E.ら,Annals of transplantation 2011 16,59-71)。これらの細胞は、成体幹細胞の集団であるか、単に用いる技術による人為的影響であるかのいずれかであると仮定されてきた。いずれの場合も、このような細胞はまれであり、研究および治療を目的とする多能性細胞の十分な供給源にはならない。
【発明の概要】
【0005】
本明細書には、国際公開第2013/163296号およびObokataら,Nature 2014 505:641-647(それぞれ参照により本明細書に組み込まれる)に開示されている方法よりも効率、収率および/または品質が向上するよう改善された、多能性細胞、例えばSTAP細胞の生成方法が記載される。本明細書にはこのほか、本発明の方法により生成した細胞に関連する方法および使用が記載される。
【図面の簡単な説明】
【0006】
図1A~図1Dは、CD45陽性体細胞からのOct4発現細胞生成を示す図である。図1Aはストレス処理胞のOct4-GFP発現を示す。ストレス処理細胞がOct4-GFPを発現するのに対し、未処理対照は発現しなかった。ストレス処理群の右上にはOct4発現コロニーの拡大画像が示されている。スケールバーは100μmを表す。
図1A~図1Dは、CD45陽性体細胞からのOct4発現細胞生成を示す図である。図1Bはストレス処理細胞および未ストレス処理対照の集団解析を示す。ストレス処理群にのみ第5日にGFP発現細胞集団が観察される。
図1A~図1Dは、CD45陽性体細胞からのOct4発現細胞生成を示す図である。図1Cは、ストレス処理前およびストレス処理後第7日のCD45陽性細胞の細胞サイズ解析を示す。
図1A~図1Dは、CD45陽性体細胞からのOct4発現細胞生成を示す図である。図1Dは、ストレス処理後のCD45陽性細胞の経時的変化を示す。
図2A~図2Bは、動物カルス細胞(ACC)の特徴付けを示す図である。図2Aは多能性マーカー遺伝子の経時的な発現の変化を示す。メッセンジャーRNAレベルをGAPDHに対して正規化した(n=3、平均+S.D.)。
図2A~図2Bは、動物カルス細胞(ACC)の特徴付けを示す図である。図2BはOct4プロモーターおよびNanogプロモーター遺伝子のメチル化解析を示す。
図3A~図3Dは、ストレス処理後の細胞改変を示す図である。図3Aは、ACC生成段階におけるストレス防御遺伝子の相対遺伝子発現量を示す。第3日および第7日に試料を収集し、CD45陽性細胞と比較した(n=3、平均+S.D.)。図3Bは総細胞内ATP測定値を示す(n=3、平均+S.D.)。図3CはROS測定値を示す。エラーバーはSDを表す。図3DはmtDNA複製因子の相対遺伝子発現量を示す(n=3、平均+S.D.)。
図3A~図3Dは、ストレス処理後の細胞改変を示す図である。図3Aは、ACC生成段階におけるストレス防御遺伝子の相対遺伝子発現量を示す。第3日および第7日に試料を収集し、CD45陽性細胞と比較した(n=3、平均+S.D.)。図3Bは総細胞内ATP測定値を示す(n=3、平均+S.D.)。図3CはROS測定値を示す。エラーバーはSDを表す。図3DはmtDNA複製因子の相対遺伝子発現量を示す(n=3、平均+S.D.)。
図4A~図4Bは、ACCからのキメラマウス形成を示す図である。図4Aはキメラマウス形成のスキームを示す。パネル(i)は、トリプシンによりACを単一細胞に分離した後または(パネルii)ACを小片に切り分けた後に胚盤胞に注入したことを示す。
図4A~図4Bは、ACCからのキメラマウス形成を示す図である。図4Bはキメラ寄与解析を示す。9個体の仔の組織をFACSにより解析した。
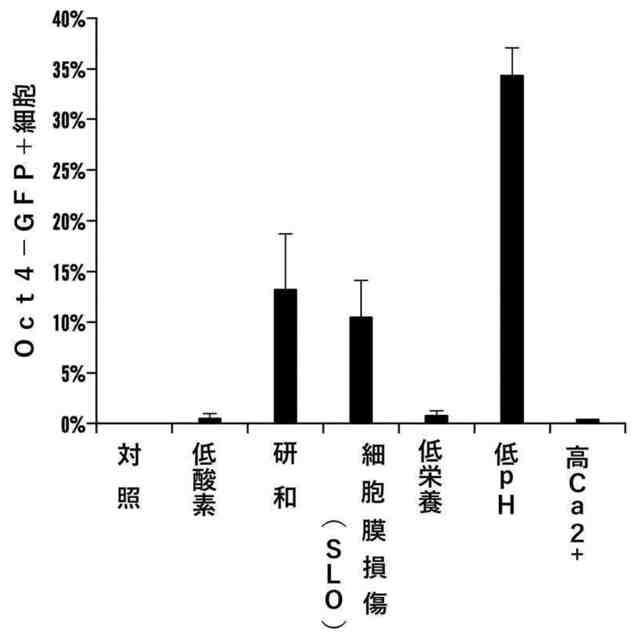
図5A~図5Cは、ACC生成条件を用いた実験を示す図である。図5Aは、CD45陽性細胞を様々なストレスに曝露し、FACSによりOct4-GFP発現を解析したことを示す。ストレス処理後の生存細胞に含まれるOct4-GFP発現細胞の百分率(n=3、平均+S.D.)。
図5A~図5Cは、ACC生成条件を用いた実験を示す図である。図5BはpH条件の決定を示す。CD45陽性細胞を様々なpHの溶液に曝露した。ストレス処理から3日後、FACSによりOct4-GFP発現を解析した。
図5A~図5Cは、ACC生成条件を用いた実験を示す図である。図5Cは培養条件の決定を示す。ストレス処理細胞を様々な培地で培養した。第14日、GFP発現ACの数をカウントした(n=3、平均+S.D.)。
図6A~図6Bは、ICRマウス由来のCD45陽性細胞からのACC生成を示す図である。図6Aはストレス処理後のCD45陽性細胞の経時的変化を示す。FACSによりE-カドヘリンおよびSSEA-1の発現を解析した。図6Bは、RT-PCRによりE-カドヘリン/SSEA1二重陽性細胞のOct4遺伝子発現を確認したことを示す(n=3、平均+S.D.)。
図7A~図7Bは、GOFマウス由来の様々な組織からのACC生成を示す図である。図7Aはストレス処理後のOct4-GFP発現細胞の割合を示す。様々な組織から体細胞を単離し、様々なストレスに曝露した。FACSによりOct4-GFP発現を解析した。
図7A~図7Bは、GOFマウス由来の様々な組織からのACC生成を示す図である。図7Bは、様々な組織に由来するACCの胚性遺伝子発現を示す。遺伝子発現をGAPDHにより正規化した(n=3、平均+S.D.)。
最初の7日間のストレス防御遺伝子の相対遺伝子発現量を示す図である。ストレス処理後、第1日、第3日および第7日に細胞を収集し、遺伝子発現を天然のCD45陽性細胞と比較した。青のグラフは熱ショックタンパク質の遺伝子発現を表す。緑のグラフはDNA修復遺伝子の発現を表す。赤のグラフは酸化還元遺伝子の発現を表す。Y軸は発現量の相対倍数を表す。
ACCの分化を示す図である。グラフはキメラ寄与解析を示す。様々な体細胞に由来するACCを用いて形成されたキメラ胚をFACSにより解析した。グラフはE13.5~E15.5のキメラ胚5例の平均を示す。
ストレス処理が間葉-上皮転換(MET)を介して体細胞のリプログラミングを引き起こしたことを示す図である。天然の細胞ならびにストレス処理開始後3日目および7日目の細胞にMET関連遺伝子の発現がみられる。y軸は、その遺伝子の発現レベルを用いて試料中のレベルに対して正規化した%発現を示す。
ストレス前およびストレス後の細胞集団のFACS解析を示す図である。GFP発現は明らかであり、各被験組織型由来のストレス後細胞集団に多能性細胞が生成されたことを示している。
図12A~図12Eは、低pH処理が運命拘束された体細胞に運命転換を誘導したことを示す図である。図12Aは実験プロトコルの模式図を示す。図12Bはフローサイトメトリー解析(上段:oct3/4::GFP
+
/CD45
-
;下段:未処理CD45
+
細胞)を示す。y軸はOct3/4:GFP細胞の数であり、X軸はCD45
+
細胞の数である。両軸とも主要単位である0、100、1000および10,000が記されている。図12Cは、培養中の経時的なoct3/4::GFP
+
生存細胞数およびoct3/4::GFP
-
生存細胞数のグラフを示す。図12Dは、Oct3/4::GFP
+
細胞(左側のピーク)およびCD45
+
細胞(右側のピーク)の細胞の大きさのグラフを示す。図12Eは、単離oct3/4::GFP
+
スフィアのtcrβのゲノム再編成をゲノムPCRにより解析した結果を示す。
図12A~図12Eは、低pH処理が運命拘束された体細胞に運命転換を誘導したことを示す図である。図12Aは実験プロトコルの模式図を示す。図12Bはフローサイトメトリー解析(上段:oct3/4::GFP
+
/CD45
-
;下段:未処理CD45
+
細胞)を示す。y軸はOct3/4:GFP細胞の数であり、X軸はCD45
+
細胞の数である。両軸とも主要単位である0、100、1000および10,000が記されている。図12Cは、培養中の経時的なoct3/4::GFP
+
生存細胞数およびoct3/4::GFP
-
生存細胞数のグラフを示す。図12Dは、Oct3/4::GFP
+
細胞(左側のピーク)およびCD45
+
細胞(右側のピーク)の細胞の大きさのグラフを示す。図12Eは、単離oct3/4::GFP
+
スフィアのtcrβのゲノム再編成をゲノムPCRにより解析した結果を示す。
図12A~図12Eは、低pH処理が運命拘束された体細胞に運命転換を誘導したことを示す図である。図12Aは実験プロトコルの模式図を示す。図12Bはフローサイトメトリー解析(上段:oct3/4::GFP
+
/CD45
-
;下段:未処理CD45
+
細胞)を示す。y軸はOct3/4:GFP細胞の数であり、X軸はCD45
+
細胞の数である。両軸とも主要単位である0、100、1000および10,000が記されている。図12Cは、培養中の経時的なoct3/4::GFP
+
生存細胞数およびoct3/4::GFP
-
生存細胞数のグラフを示す。図12Dは、Oct3/4::GFP
+
細胞(左側のピーク)およびCD45
+
細胞(右側のピーク)の細胞の大きさのグラフを示す。図12Eは、単離oct3/4::GFP
+
スフィアのtcrβのゲノム再編成をゲノムPCRにより解析した結果を示す。
図13A~図13Bは、低pH誘導Oct3/4
+
細胞が多能性を有することを示す図である。図13Aは、d7の低pH誘導oct3/4::GFP
+
細胞のqPCRの遺伝子発現解析をCD45
+
細胞と比較したグラフを示す(各組とも左から右に向かってoct3/4、nanog、sox2、ecat1、esg1、dax1およびklf4の発現を表す)。第3日および第7日に試料を収集し、CD45陽性細胞と比較した(n=3、平均+S.D.)。図13Bは、バイサルファイトシーケンシングの結果がoct3/4プロモーターおよびnanogプロモーター領域のものであることを示す。CD45
+
細胞は、追加の培養の有無に関係なく、両プロモーターに密にメチル化されたパターンを示した。
図14A~図14Bは、他の組織供給源からもSTAP細胞が得られることを示す図である。図14Aは、多数の組織(各組とも左から右に向かってCD45
+
細胞、骨髄、脳、肺、筋肉、脂肪、線維芽細胞、肝臓および軟骨細胞を表す)の培養d7のoct3/4::GFP
+
細胞生成の割合を示す。図14Bは、oct3/4::GFP
+
細胞クラスターの遺伝子発現解析のグラフを示す(各組とも左から右に向かってOct3/4、Nanog、Sox2、Klf4およびRex1の発現を表す)。
図15A~図15Bは、STAP細胞の多能性細胞としての特徴付けを示す図である。図15AはSTAP細胞におけるES細胞マーカーの遺伝子発現のグラフを示す(各組とも左から右に向かってES、EpiSC、STAPおよびCD45を表す)。図15BはSTAP細胞におけるX染色体不活化の%のグラフを示す。
様々なストレスに曝露したCD45陽性細胞についてFACSにより分析したOct4-GFP発現のグラフを示す図である。ストレス処理後の生存細胞におけるOct4-GFP発現細胞の百分率(n=3、平均+S.D.)。
pH条件決定のグラフを示す図である。CD45陽性細胞を様々なpHの溶液に曝露した。ストレス処理から3日後、FACSによりOct4-GFP発現を解析した(n=3、平均+S.D.)。
培養条件決定のグラフを示す図である。ストレス処理細胞を様々な培地で培養した。第14日、GFPを発現するストレス変化細胞塊の数をカウントした(n=3、平均+S.D.)。
図17A~図17Bは、ICRマウス由来のCD45陽性細胞からのSAC生成を示す図である。図17Aはストレス処理後のCD45陽性細胞の経時的変化を示す。FACSによりE-カドヘリンおよびSSEA-1の発現を解析した。図17Bは、RT-PCRによって確認したE-カドヘリン/SSEA1二重陽性細胞のOct4遺伝子発現のグラフを示す(n=3、平均+S.D.)。
図18A~図18Bは、GOFマウス由来の様々な組織からのSAC生成を示す図である。図18Aはストレス処理後のOct4-GFP発現細胞の割合のグラフを示す。様々な組織から体細胞を単離し、様々なストレスに曝露した。FACSによりOct4-GFP発現を解析した。各組とも左から右に向かってBM、脳、肺、筋肉、脂肪、線維芽細胞および肝臓を表す。
図18A~図18Bは、GOFマウス由来の様々な組織からのSAC生成を示す図である。図18Bは、様々な組織に由来するSACの胚性遺伝子発現のグラフを示す。GAPDHにより遺伝子発現を正規化した(n=3、平均+S.D.)。各組とも左から右に向かってOct4、Nanog、Sox2、Klf4およびEcat1を表す。
最初の7日間のストレス防御遺伝子の相対遺伝子発現量のグラフを示す図である。ストレス処理後、第1日、第3日および第7日に細胞を収集し、遺伝子発現を天然のCD45陽性細胞と比較した。Y軸は発現量の相対倍数を表す。
SACおよびCD45+細胞由来のSACから誘導されたキメラマウスのTCRβ鎖再編成解析を示す図である。2Nキメラマウス#1、#2、#3、#5、#6、#7、#8および#9に再編成DNAの発現がみられた。
4Nキメラマウスのジェノタイピング解析を示す図である。ジェノタイピングを実施したところ、129/Sv×B6GFP F1由来のSACを用いて形成した4NキメラマウスおよびICR由来の4N胚盤胞がSAC(129/Sv×B6GFP)特異的遺伝子を発現することが明らかになった。
STAP細胞がin vivoで胚組織および胎盤組織に寄与することを示す図である。グラフは、注入した細胞が胚部分にのみ寄与した胎児ならびに胚部分に加えて胎盤および卵黄嚢組織にも寄与した胎児の割合を示す。
図23A~図23Cは、FGF4処理がSTAP細胞に一部のトロホブラスト系列の特徴を誘導することを示す図である。図23Aは、STAP細胞からTS様(F4I)細胞を誘導するためのFGF4処理の模式図を示す。図23Bはマーカー発現のqPCR解析のグラフを示す。図23CはFACS解析による胎盤寄与の定量化のグラフを示す。F4I細胞とは異なり、ES細胞は検出可能なレベルで胎盤組織に寄与することはなかった。
図24A~図24Dは、STAP細胞からES細胞様幹細胞が誘導され得ることを示す図である。図24AはSTAP細胞からの幹細胞誘導の模式図を示す。図24Bは、STAP-S細胞が120日間にわたる維持培養で安定して増殖したことを示す。同様の結果が16種類の独立した系統で得られた。これに対し、親STAP細胞では急激に数が減少した。図24Cはマーカー遺伝子発現のqPCR解析のグラフを示す。ES細胞およびSTAP-S細胞は、CD45
+
細胞には発現しなかった多能性関連遺伝子を発現した。図24Dは、バイサルフェートシーケンシングによるDNAメチル化試験の略図を示す。
図24A~図24Dは、STAP細胞からES細胞様幹細胞が誘導され得ることを示す図である。図24AはSTAP細胞からの幹細胞誘導の模式図を示す。図24Bは、STAP-S細胞が120日間にわたる維持培養で安定して増殖したことを示す。同様の結果が16種類の独立した系統で得られた。これに対し、親STAP細胞では急激に数が減少した。図24Cはマーカー遺伝子発現のqPCR解析のグラフを示す。ES細胞およびSTAP-S細胞は、CD45
+
細胞には発現しなかった多能性関連遺伝子を発現した。図24Dは、バイサルフェートシーケンシングによるDNAメチル化試験の略図を示す。
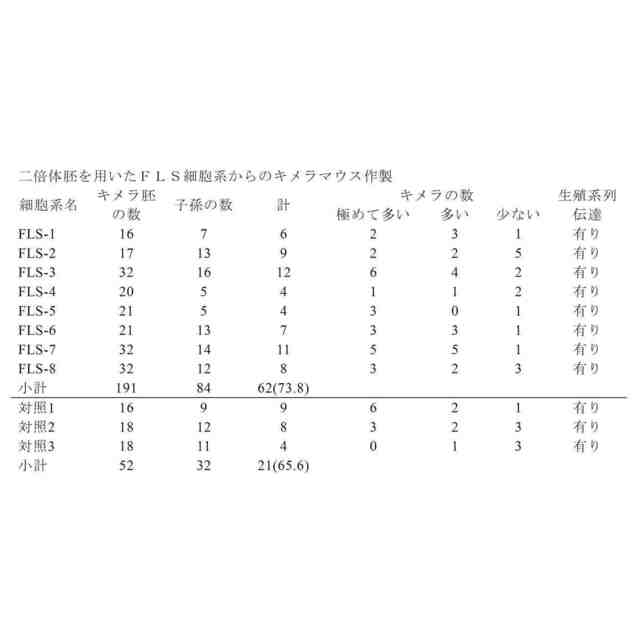
図25A~図25Bは、STAP幹細胞が多能性であり、生殖系列伝達および四倍体補完法に適合性があることを示す図である。図25Aは、胚盤胞注入アッセイ(2N)におけるキメラマウスの様々な組織へのSTAPS細胞の寄与率のグラフを示す。図25Bは胎盤組織への寄与率のグラフを示す。親STAP細胞およびTS細胞とは異なり、STAPS細胞は胎盤寄与の能力をもはや保持していなかった。3種類の独立した系統を試験し、いずれも胚部分への実質的な寄与を示した。
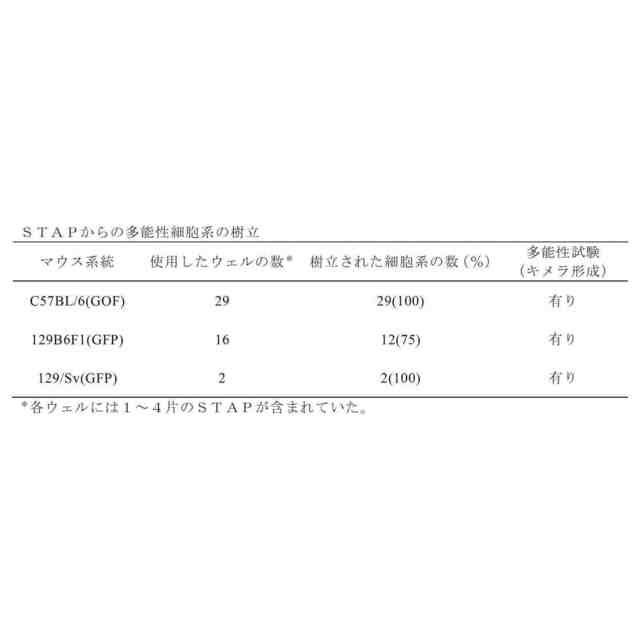
酸洗浄により誘導された多能性を示す図である。
上:髄腔内SSP-SAPで処置したラットではカプサイシン注射後の肢引っ込め閾値の低下によって示される機械的痛覚過敏が減少することを示す図である。差が最も大きくなるカプサイシン注射10分後の反応をのちのグラフに示す。下:脊髄幹細胞を埋植してから5週間後、カプサイシン誘導性痛覚過敏が回復することを示す図である。
最初に、痛覚過敏状態を大幅に減少させるSSP-SAPをi.t.注射したラットの足にカプサイシンを注射し(図27を参照されたい)、次いで、幹細胞の腰髄i.t.注射により処置した後の触覚反応(上)および熱反応(下)を示す図である。「元の反応」は何らかの処置を実施する前のカプサイシンに対する痛覚過敏反応を示す。「BL1」は、2週間前にSSP-SAPまたは不活性なBlank-SAPのいずれかで処置したラットにカプサイシン注射を実施する前のベースラインの反応を示す。「BL2」は、カプサイシン注射を実施せずに幹細胞を送達してから1~2日後のベースラインの反応を示す。幹細胞埋植物によりSSP-SAP処置ラットの痛覚過敏反応を未処置ラットおよびBlank-SAP処置対照のものに戻すことが可能であることに注目されたい。
幹細胞埋植物によってカプサイシン感受性が回復したラットでは、NK1-Rの特異的アンタゴニストの効力が増大することを示す図である。未処置ラット(○、□;左パネル;およびBlank-SAPを投与した後、幹細胞を送達したラット、不掲載)では、両型の痛覚過敏ともL-733,060のIC50が約0.3mM(30μLのi.t.注射)であるのに対し、幹細胞回復ラット(右パネル)では、触覚痛覚過敏(■)のIC50が約30μM、熱痛覚過敏(●)のIC50が約5μMである。
【発明を実施するための形態】
【0007】
(詳細な説明)
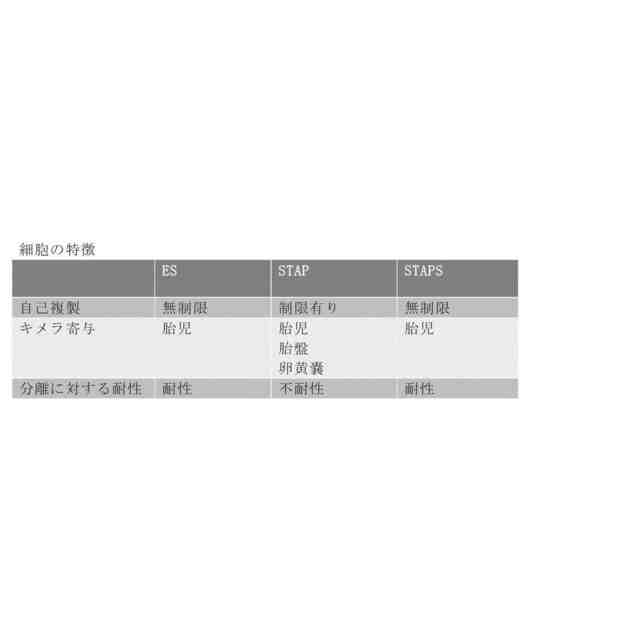
本明細書に記載される技術の諸態様は、細胞からの多能性細胞の作製または生成に関する。本明細書に記載される技術の諸態様は、ストレスが、細胞に外来遺伝子も、転写産物も、タンパク質も、核成分も、細胞質も導入することを必要とせず、また細胞融合も必要とせずに細胞からの多能性幹細胞の産生を誘導し得るという本発明者らの発見に基づくものである。いくつかの実施形態では、ストレスが細胞の細胞質および/またはミトコンドリアの量の減少を誘導し、脱分化過程を惹起して多能性細胞を生じさせる。いくつかの実施形態では、ストレスが、例えばストレスに曝露した細胞の少なくとも10%に細胞膜の破壊を引き起こす。これらの多能性細胞は、三胚葉のそれぞれに分化する能力(in vitroおよび/またはin vivo)、in vivoでの奇形腫様細胞塊の生成ならびに生きた胚および/またはキメラマウスを生成する能力のうちの1つまたは複数を特徴とする。
【0008】
本明細書には、特に限定されないが細胞の細胞質および/またはミトコンドリアの量を減少させるストレスを含めた特定の環境ストレスで細胞を処理すると、ミトコンドリアの活性が低下し、脱分化に関連するゲノム領域が脱メチル化され、細胞が既知の脱分化経路のマーカーを示し得ることを示す実験が記載される。したがって、いくつかの実施形態では、細胞から多能性細胞を生成する方法が本明細書に提供され、この方法は、細胞から細胞質および/またはミトコンドリアの少なくとも約40%を除去することと、多能性細胞または多能性マーカーを示す細胞を選択することとを含み、細胞が組織中に存在しない。本明細書にはこのほか、細胞から多能性細胞を生成させ得るその他のストレス処理が記載される。
【0009】
便宜上、本願の明細書、実施例および添付の特許請求の範囲に使用する特定の用語をここにまとめる。別途記載されるか、文脈から読み取れない限り、以下の用語および語句は以下に記載する意味を包含する。別途明記されるか、文脈から明らかでない限り、以下の用語および語句は、それと関係のある技術分野で付与されている意味を排除するものではない。定義は、特定の実施形態の説明に役立つよう記載されるものであり、また本発明の範囲は請求項によってのみ限定されることから、特許請求される発明を限定することを意図するものではない。特に明記されない限り、本明細書で使用される技術用語および科学用語はいずれも、本発明が属する技術分野の当業者によって一般に理解されるものと同じ意味を有する。
【0010】
本明細書で使用される「含む」という用語は、組成物、方法およびその方法または組成物に必須の各構成要素(1つまたは複数)を指すのに使用されるが、必須であるかどうかに関係なく、明記されていない要素を包含する可能性もある。
(【0011】以降は省略されています)
この特許をJ-PlatPat(特許庁公式サイト)で参照する
関連特許
他の特許を見る






 特許ウォッチ
特許ウォッチ