TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025093955
公報種別
公開特許公報(A)
公開日
2025-06-24
出願番号
2025026896,2021563374
出願日
2025-02-21,2020-05-29
発明の名称
単為生殖のための遺伝子
出願人
キージーン ナムローゼ フェンノートシャップ
代理人
個人
,
個人
,
個人
,
個人
主分類
C12N
15/29 20060101AFI20250617BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】作物においてアポミクシス(無融合種子形成(agamospermy)とも呼ばれる)を誘導するための手順、およびアポミクティック植物及びアポミクティック種子を生産するための方法を提供する。また、アポミクシスのプロセス、特に単為生殖に関与する遺伝子及びタンパク質を提供する。
【解決手段】本発明は、タンポポ属(Taraxacum)の単為生殖遺伝子のヌクレオチド配列及びアミノ酸配列、並びにそれらの(機能的)相同体、断片及びバリアントを提供し、これらは、アポミクシスの一部として単為生殖を実現する。また、単為生殖植物及びこれらを作製する方法が提供され、分子マーカー及びこれらを使用する方法も提供される。
【選択図】なし
特許請求の範囲
【請求項1】
a)配列番号1、6又は11のアミノ酸配列を有するタンパク質をコードする遺伝子;
b)配列番号2、7又は12のヌクレオチド配列を有するプロモーター;
c)配列番号3、8又は13のヌクレオチド配列を有するコード配列;
d)配列番号4、9又は14のヌクレオチド配列を有する3’UTR;
e)配列番号5、10又は15のヌクレオチド配列を有する遺伝子;
f)a)~e)うちのいずれか1つのバリアント又は断片
のうちの少なくとも1つを含む、植物における単為生殖に関連する核酸であって、
好ましくは単為生殖において機能的である
核酸。
続きを表示(約 1,000 文字)
【請求項2】
キメラ遺伝子、遺伝子構築物又は核酸ベクターに含まれる、請求項1に記載の核酸。
【請求項3】
a)請求項1に記載の核酸によってコードされる;
b)配列番号1、6若しくは11のアミノ酸配列を有する;並びに/又は
c)a)及び/若しくはb)のバリアント若しくは断片である、
植物における単為生殖に関連するタンパク質であって、
好ましくは単為生殖において機能的である
タンパク質。
【請求項4】
広義のセイヨウタンポポ(Taraxacum officinale)種ではない植物又は植物細胞であって、請求項1に記載の核酸及び/又は請求項3に記載のタンパク質を含み、好ましくは、アブラナ科(Brassicaceae)、ウリ科(Cucurbitaceae)、マメ科(Fabaceae)、イネ科(Gramineae)、ナス科(Solanaceae)、キク科(Asteraceae(コンポジテ(Compositae)))、バラ科(Rosaceae)及びイネ科(Poaceae)からなる群から選択される科由来である、植物又は植物細胞。
【請求項5】
遺伝子改変によって又は遺伝子移入によって請求項1に記載の核酸を含み、好ましくは前記核酸が前記植物又は植物細胞のゲノムに統合されている、請求項4に記載の植物又は植物細胞。
【請求項6】
単為生殖が可能である、請求項4又は5に記載の植物又は植物細胞。
【請求項7】
さらにアポマイオシスが可能であり、好ましくはアポミクシスが可能である、請求項4~6のいずれか一項に記載の植物又は植物細胞。
【請求項8】
請求項4~7のいずれか一項に記載の植物又は植物細胞の種子、植物部分又は植物産物。
【請求項9】
a)単為生殖を誘導することが可能である請求項1に記載の核酸を1つ又は複数の植物細胞に導入するステップと;
b)前記核酸を含む植物細胞を選別するステップであって、好ましくは、前記核酸が前記植物細胞のゲノムに統合されているステップと;
c)前記植物細胞から植物を再生するステップと
を含む、単為生殖植物を作出する方法。
【請求項10】
請求項9に記載のステップa)~c)を含み、ステップa)の前記1つ又は複数の植物細胞はアポマイオシスが可能である、アポミクティック植物を作出する方法。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本発明はバイオテクノロジーの分野に、特に、植物育種を含む植物バイオテクノロジーに関する。本発明は、例えばアポミクシス及び単数体誘導に関連し且つこれらにおいて有用な遺伝子の同定及び使用に特に関する。本発明は、単為生殖に関連する遺伝子、並びにそのコードされたタンパク質及び双方の断片に特に関する。本発明は、植物及び作物において単為生殖を抑制及び/又は誘導する方法に、アポマイオシス性遺伝子(複数可)と特に組み合わせたアポミクシスのための、又はその染色体を倍加して倍加単数体を生産することができる単数体植物の作出のための、遺伝子及び/若しくはタンパク質又はそれらの断片の使用に、さらに関する。
続きを表示(約 4,300 文字)
【背景技術】
【0002】
アポミクシス(無融合種子形成(agamospermy)とも呼ばれる)とは、種子を通しての無性植物生殖である。アポミクシスはおよそ400種の顕花植物種で報告されている(Bicknell及びKoltunow、2004)。顕花植物のアポミクシスは以下の2つの形態で生じる:
(1)配偶体のアポミクシス、この場合、胚は、単為生殖によって、非減数未受精卵細胞から発生する;
(2)胞子体アポミクシス、この場合、胚は、胞子体細胞から体細胞性に発生する。
配偶体アポミクトの例は、タンポポ(タンポポ属の種(Taraxacum sp.))、ヤナギタンポポ(ヤナギタンポポ属の種(Hieracium sp.))、ケンタッキーブルーグラス(ナガハグサ(Poa pratensis))及びイースタンガマグラス(トリプサクム・ダクチロイデス(Tripsacum dactyloides))である。胞子体アポミクシスの例は、シトラス(シトラス属の種(Citrus sp.))及びマンゴスチン(ガルシニア・マンゴスタナ(Garcinia mangostana))である。配偶体のアポミクシスは、2つの発生プロセス:
(1)減数分裂組換え及び減数の回避(アポマイオシス);及び
(2)受精なしでの卵細胞の胚への発生(単為生殖)
が、関与する。
【0003】
アポミクティック的に生産された種子は、親植物と遺伝的に同一である。アポミクシスは植物育種に極めて有用であり得ることはずっと以前から認識されていた(Asker、1979;Hermsen、1980;Asker及びJerling、1990;Vielle-Calzadaら、1995)。作物へのアポミクシスの導入に関して最も明白な利点とは、ヘテロ強勢F1雑種の純粋な育種である。ほとんどの作物では、F1雑種は優良の品種である。しかし、有性作物では、F1雑種の自家受精は、F2後代植物のゲノムにおける組換えによるヘテロ強勢の喪失を引き起こすので、F1雑種は、再度、同系交配のホモ接合型の両親の交配によって各世代で作出する必要がある。有性F1種子を生産することは、何度も繰り返される、複雑な、費用のかかるプロセスである。対照的に、アポミクティックF1雑種は、永続的に純粋種を生み出すと思われる。換言すれば、種子を通してのF1雑種の遺伝的固定及び均一な後代植物の作出が可能となる。
【0004】
アポミクシスによるF1固定とは、その遺伝的複雑さが何であれ、任意の遺伝子型が一段階で純粋種を生み出すことになる、アポミクシスの一般的特性の特例である。このことが意味するところは、アポミクシスを使用して、多遺伝子性の定量的形質を即座固定することが可能であることである。ほとんどの収量形質は多遺伝子性であることに留意されたい。アポミクシスは、多重形質(例えば、種々の耐性、いくつかの導入遺伝子、又は多重量的形質遺伝子座)のスタッキング(又はピラミッド化)に使用可能である。アポミクシスを使用しない場合、そのような一連の形質を固定するためには、各形質の遺伝子座を個別にホモ接合型とし、後に組み合わせる必要がある。形質に関与する遺伝子座の数が増加するにつれて、交配によってこれらの形質遺伝子座をホモ接合型にすることは、時間がかかり、事業遂行上難しい課題となり、それによって費用がかかるようになる。さらに、対立遺伝子間の特定のエピスタシス相互作用は、ホモ接合性によって喪失する。アポミクシスを使用すると、このタイプの非相加的な遺伝的変異を固定することが可能になる。したがって、アポミクシス、種子を通してのクローン繁殖は、植物育種、商業的種子生産及び農業におけるパラダイムシフトを引き起こす可能性を有する(van Dijkら 2016、Van Dijk及びSchauer 2016)。
【0005】
任意の遺伝子型を即座に固定することに加えて、その複雑性が何であれ、アポミクシスのさらなる重要な農業用途が存在する。有性種間雑種及び同質倍数体については、減数分裂の問題に起因する不稔に悩まされている場合が多い。アポミクシスは減数分裂をスキップするので、アポミクシスを使用すると、種間雑種と同質倍数体に関するそういった不稔の問題を解決できる。アポミクシスは雌性ハイブリダイゼーションを防止するので、雄性不稔と組み合わせたアポミクシスは、トランスジェニック作物の野生関連種における導入遺伝子の遺伝子移入を防止する、導入遺伝子の封じ込め向けに提案された(Daniell、2002)。虫媒受粉作物(例えば、アブラナ属(Brassica))では、アポミクティック種子結実は、不十分な受粉媒介者のサービスによって制限されないはずである。これは、受粉ハチの集団の増大する健康問題(ミツバチヘギイタダニ(Varroa mite)感染症、アフリカ系キラービー等)と照らして、より重要となってきている。ジャガイモのような塊茎増殖作物では、アポミクシスは、クローン的に優れた遺伝子型を維持するが、クリーンな生産、封じ込め及び認証におけるウイルス伝染及び関連するコストの現在のリスクを軽減又は排除をもすると思われる。また、アポミクティック種子の貯蔵コストは、塊茎又はその他の栄養繁殖した植物部分の貯蔵コストよりもはるかに低い。観賞用では、アポミクシスは労働集約的で費用のかかる組織培養増殖に取って代わる可能性がある。一般に、アポミクシスは栽培品種の開発と植物増殖のコストを強力に削減すると考えられる。
【0006】
残念なことに、アポミクシスは主たる作物のいずれでも存在しない。有性作物にアポミクシスを導入する試みは数多くある。例をあげると、アポミクシス遺伝子の遺伝子移入、有性モデル種の突然変異、ハイブリダイゼーションによるアポミクシスのデノボ生産、及び候補遺伝子のクローニング。遠縁交配による野生アポミクトから作物種へのアポミクシス遺伝子の遺伝子移入は、これまで成功していない(例えば、トリプサクム・ダクチロイデスからトウモロコシへのアポミクシス - Savidan,Y.、2001;Morganら、1998;国際公開第97/10704号)。突然変異有性モデル種に関して、国際公開第2007/066214号では、シロイヌナズナ属(Arabidopsis)においてDyadと呼ばれるアポマイオシス変異体の使用が記載されている。しかし、Dyadは浸透率が極めて低い劣性突然変異である。作物種では、この突然変異の用途は限定される。2つの有性エコタイプ間のハイブリダイゼーションによるデノボでのアポミクシスの生産は、農業上興味深いアポミクトをもたらさなかった(米国特許出願公開第2004/0168216A1号及び米国特許出願公開第2005/0155111A1号)。トウモロコシにおけるトランスポゾンタグ付けによる候補アポミクシス遺伝子のクローニングが、米国特許出願公開第2004/0148667号に記載されている。アポミクシスを誘導すると仮定される、伸長遺伝子のオルソログが特許請求された。しかし、Barrell及びGrossniklaus(2005)によると、伸長遺伝子は減数分裂IIをスキップし、したがって母親の遺伝子型を維持せず、このことによって、その伸長遺伝子は有用性がかなり低いものとなる。
【0007】
米国特許出願公開第2006/0179498号では、いわゆる逆育種がアポミクシスの代替となると思われることが記載された。しかし、逆育種は、技術的に複雑なin vitroの実験室手順であり、一方、アポミクシスは植物自体によって実行されるin vivo手順である。さらに、逆育種を使用する場合、いったん親系統が再構築されると(倍加配偶子ホモ接合体)、交配をなおも実行する必要がある。
【0008】
天然アポミクトにおけるアポミクシスは一般に遺伝的ベースを有する(Ozias-Akins及びVan Dijkによるレビュー、2007)。したがって、代替法は、天然アポミクティック種からのアポミクシス遺伝子の単離となり得る。しかし、これは容易なタスクではない、その理由は、天然アポミクトは倍数体ゲノムを有することが多く、倍数体でのポジショナルクローニングは極めて困難である。他の複雑な要因とは、アポミクシスに特異的な染色体領域における組換えの抑制、反復配列であり、交配における分離の歪みである。
【発明の概要】
【0009】
本明細書に記載のように、現在の技術水準の限界の少なくとも一部がない、作物においてアポミクシスを誘導するための手順に対するニーズがある。特に、アポミクティック植物及びアポミクティック種子を生産するための方法に対するニーズがある。また、アポミクシスのプロセス、特に単為生殖に関与する遺伝子及びタンパク質を提供するニーズもあり、これら遺伝子及びタンパク質は、作物にアポミクシスを導入する際に使用するのに適しているとともに、アポミクティック経路を実質的に模倣することができる。
【0010】
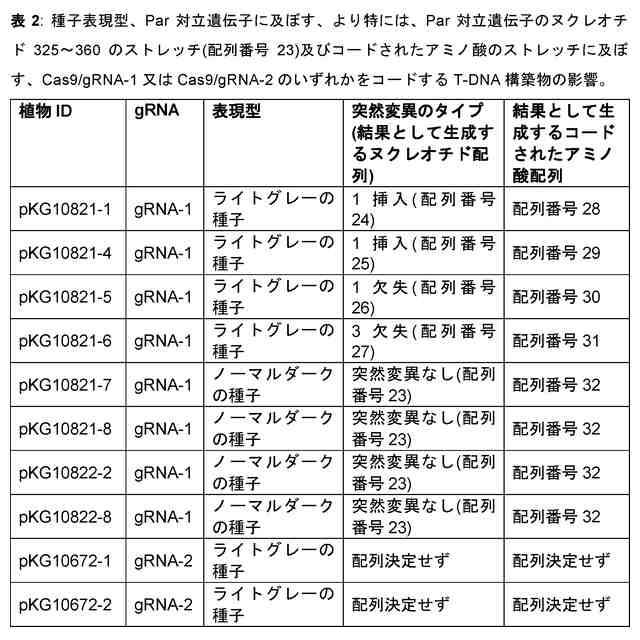
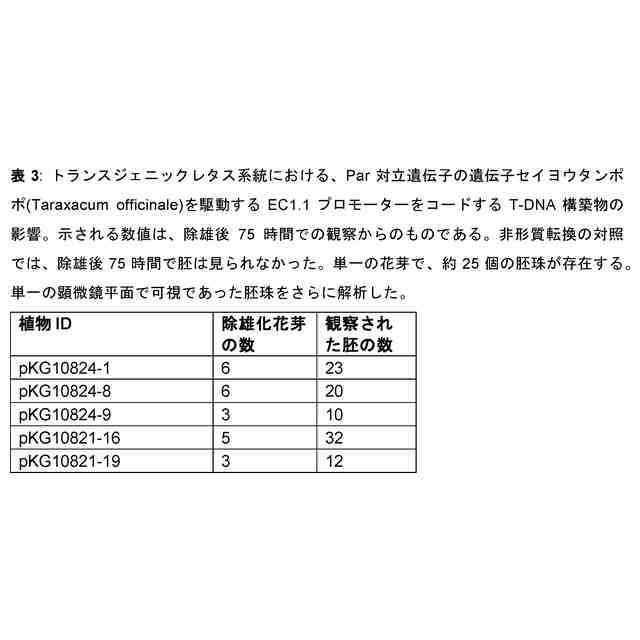
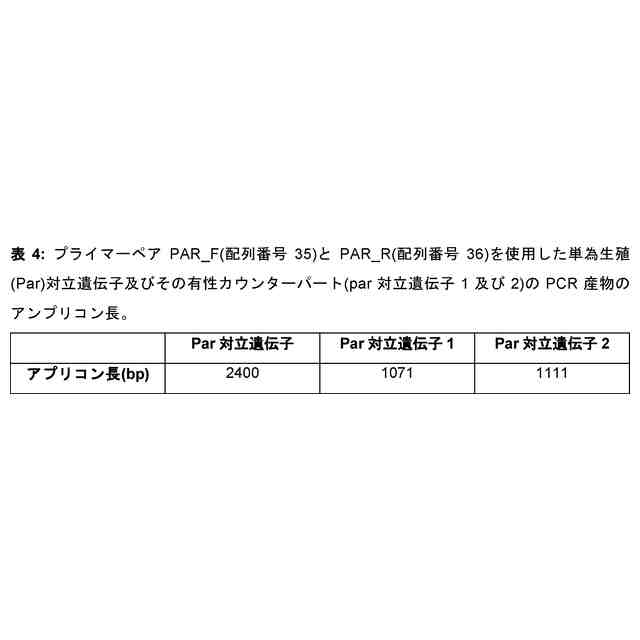
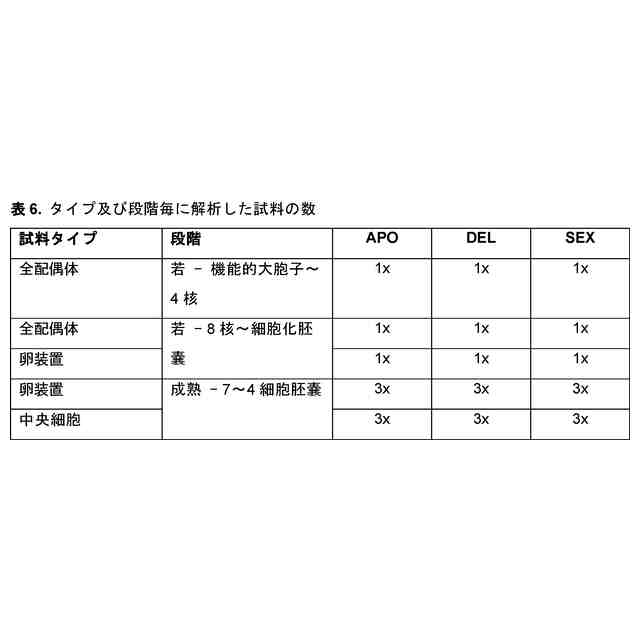
本発明者らは今回、単為生殖遺伝子座及び遺伝子、単為生殖表現型に関連する対立遺伝子(本明細書では単為生殖対立遺伝子又はPar対立遺伝子として指示される)及び非単為生殖表現型(本明細書では有性若しくは非単為生殖対立遺伝子又はpar対立遺伝子と指示される)、それらの遺伝子配列、すなわちプロモーター又は5’UTR配列、コード配列、3’UTR配列並びにコードされたタンパク質配列を同定且つ単離した。単為生殖は、おそらくランダム又は標的突然変異誘発によって、形質転換によって、又は体細胞ハイブリダイゼーションによって、有性植物に直接導入することができる。有性植物の単為生殖遺伝子座の有性対立遺伝子を遺伝的に改変することによって、例えば、突然変異誘発、遺伝子導入により、又は特定の部位での二本鎖切断の導入及び相同組換えを介した挿入により、Par対立遺伝子を導入されてもよく、植物及び/又はその子孫が卵細胞を胚へと発生させることが可能となってもよい。
【図面の簡単な説明】
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
他の特許を見る




 特許ウォッチ
特許ウォッチ