TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025032175
公報種別
公開特許公報(A)
公開日
2025-03-11
出願番号
2024208739,2021549986
出願日
2024-11-29,2020-02-26
発明の名称
クラッベ病の治療に有用な組成物
出願人
ザ・トラステイーズ・オブ・ザ・ユニバーシテイ・オブ・ペンシルベニア
代理人
弁理士法人小田島特許事務所
主分類
A61K
38/47 20060101AFI20250304BHJP(医学または獣医学;衛生学)
要約
【課題】クラッベ病患者に対する改善された治療を提供する。
【解決手段】組換えアデノ随伴ウイルス(rAAV)を含む組成物であって、前記rAAVが、(a)中枢神経系の細胞を標的とするAAVカプシドと、(b)(ii)ベクターゲノムであって、(i)シグナルペプチド、および特定のアミノ酸配列を有する成熟ヒトガラクトシルセラミダーゼのタンパク質をコードし、前記タンパク質の発現を指示する調節配列の制御下にある、ガラクトシルセラミダーゼのコード配列、ならびに(ii)前記ベクターゲノムを前記AAVカプシド中にパッケージングするのに必要なAAV逆位末端配列、を含む、ベクターゲノムと、を含み、前記ベクターゲノムが、前記AAVカプシド中にパッケージングされる、組成物が提供される。
【選択図】なし
特許請求の範囲
【請求項1】
組換えアデノ随伴ウイルス(rAAV)を含む組成物であって、前記rAAVが、
(a)中枢神経系の細胞を標的とするAAVカプシドと、
(b)(ii)ベクターゲノムであって、(i)シグナルペプチド、および少なくとも配列番号10のaa43~685のアミノ酸配列を有する成熟ヒトガラクトシルセラミダーゼのタンパク質をコードし、前記タンパク質の発現を指示する調節配列の制御下にある、ガラクトシルセラミダーゼのコード配列、ならびに(ii)前記ベクターゲノムを前記AAVカプシド中にパッケージングするのに必要なAAV逆位末端配列、を含む、ベクターゲノムと、を含み、前記ベクターゲノムが、前記AAVカプシド中にパッケージングされる、組成物。
続きを表示(約 810 文字)
【請求項2】
前記AAVカプシドが、AAVhu68カプシドである、請求項1に記載の組成物。
【請求項3】
前記コード配列が、配列番号10の全長ヒトガラクトシルセラミダーゼのシグナルペプチドおよび成熟ヒトガラクトシルセラミダーゼのタンパク質(アミノ酸1~685)をコードする、請求項1または請求項2に記載の組成物。
【請求項4】
前記コード配列が、外因性ペプチドのコード配列と配列番号9のヌクレオチド127~2055の核酸配列もしくはそれと95%~99.9%同一の配列とを有するか、または配列番号9のヌクレオチド1~2055のヌクレオチド配列もしくはそれと95%~99.9%同一の配列を有する、請求項1~3のいずれか一項に記載の組成物。
【請求項5】
前記コード配列が、配列番号10の前記成熟ヒトガラクトシルセラミダーゼのタンパク質(アミノ酸43~685)および中枢神経系のヒト細胞に好適な外因性シグナルペプチドをコードする、請求項1または請求項2に記載の組成物。
【請求項6】
前記調節配列が、ベータアクチンプロモーター、イントロン、およびウサギグロビンポリAを含む、請求項1~5のいずれか一項に記載の組成物。
【請求項7】
前記調節配列が、配列番号13を含む、請求項1~6のいずれか一項に記載の組成物。
【請求項8】
前記調節配列が、配列番号15を含む、請求項1~7のいずれか一項に記載の組成物。
【請求項9】
前記調節配列が、配列番号16を含む、請求項1~8のいずれか一項に記載の組成物。
【請求項10】
前記ベクターゲノムが、配列番号19のnt198~4168の配列を有するCB7.CI.hGALC.RBGを含む、請求項1または2に記載の組成物。
(【請求項11】以降は省略されています)
発明の詳細な説明
【背景技術】
【0001】
パルボウイルスファミリーのメンバーであるアデノ随伴ウイルス(AAV)は、約4.7キロ塩基(kb)の長さの一本鎖直鎖DNA(ssDNA)ゲノムを有する小さな非エンベロープ性の正二十面体ウイルスである。野生型ゲノムは、DNA鎖の両端に逆位末端配列(inverted terminal repeats、ITR)、ならびに2つのオープンリーディングフレーム(ORF)であるrepおよびcapを含む。Repは、AAVの生活環に必要なrepタンパク質をコードする4つの重複遺伝子から構成され、capは、カプシドタンパク質の重複ヌクレオチド配列であるVP1、VP2、およびVP3を含有し、自己集合して正二十面体対称のカプシドを形成する。
続きを表示(約 4,700 文字)
【0002】
複製欠陥ヒトパルボウイルスに由来する組換えアデノ随伴ウイルス(rAAV)ベクターは、遺伝子送達に好適なビヒクルとして記載されている。典型的には、機能的なrep遺伝子およびcap遺伝子がベクターから除去されると、複製欠陥ベクターが得られる。これらの機能は、ベクター産生システムの中で提供されるが、最終ベクターには存在しない。
【0003】
これまで、ヒトまたは非ヒト霊長類(NHP)から単離されたいくつかの異なる十分に特徴付けられたAAVが存在している。異なる血清型のAAVは、異なるトランスフェクション効率を示し、異なる細胞または組織に対して指向性(tropism)を示すことが見出されている。多くの異なるAAV系統群(clade)が、WO2005/033321号に記載されており、その中には系統群Fが含まれ、AAV9、AAVhu31、およびAAVhu32という3つのメンバーのみを有することが特定されている。AAV9の構造分析は、M.A.DiMattia et al,J.Virol.(June
2012)vol.86 no.12 6947-6958.に提供されている。本論文では、AAV9が(合計)60コピーの3つの可変タンパク質(vp)を有することが報告され、それは、cap遺伝子によってコードされ、重複配列を有する。これらには、VP1(87kDa)、VP2(73kDa)、およびVP3(62kDa)が含まれ、それぞれ、1:1:10の予測された比率で存在する。VP3の配列全体は、VP2内にあり、VP2のすべては、VP1内にある。VP1は、固有のN末端ドメインを有する。精密化された座標および構造因子は、RCSB PDBデータベースから受入番号3UX1で入手可能である。
【0004】
異なる組織を脱標的化または標的化するには、いくつかの異なるAAV9バリアントが操作されている。例えば、N.Pulicheria,“Engineering Liver-detargeted AAV9 Vectors for Cardiac and Musculoskeletal Gene Transfer”,Molecular Therapy,Vol,19,no.6,p.1070-1078(June 2011)を参照されたい。血液脳関門を横切って遺伝子を送達するためのAAV9バリアントの開発も報告されている。例えば、B.E.Deverman et al,Nature Biotech,Vol.34,No.2,p204-211(published online 1 Feb 2016)and Caltech press release,A.Wetherston,www.neurology-central.com/2016/02/10/successful-delivery-of-genes-through-the-blood-brain-barrier/,accessed 10/05/2016を参照されたい。また、WO2016/0492301およびUS8,734,809も参照されたい。
【0005】
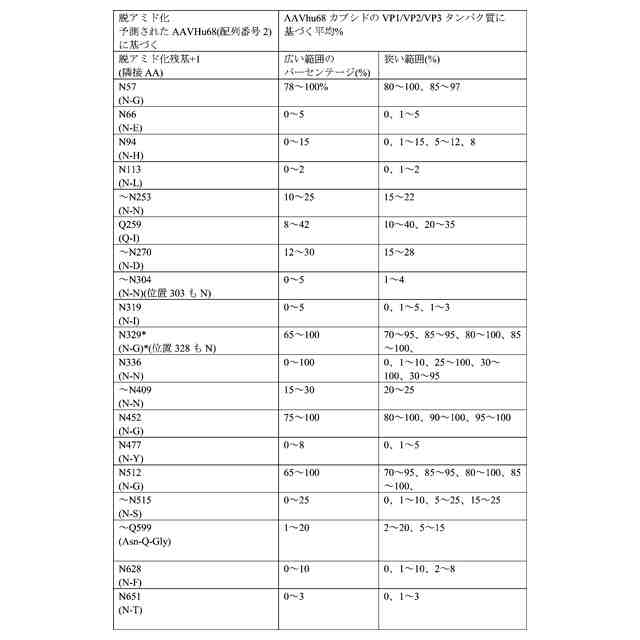
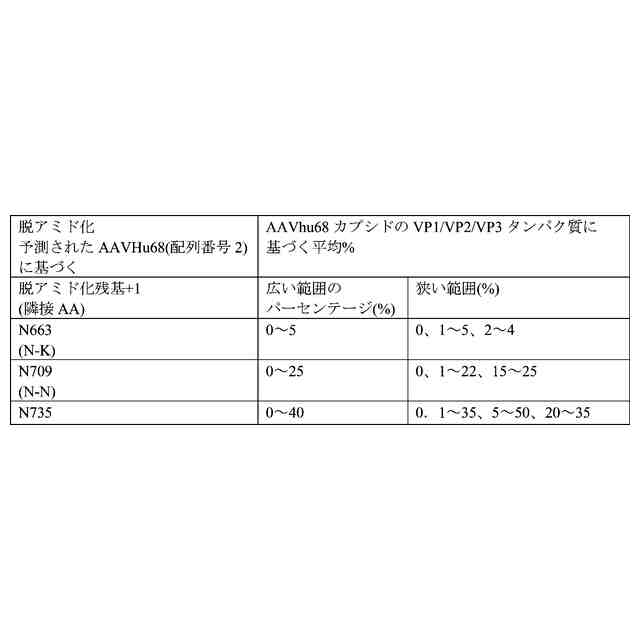
AAVhu68は、天然源由来のカプシド遺伝子の増幅後に同定され、最近、新しいAAVカプシドとして同定された。また、WO2018/160582も参照されたい。このAAVは、AAV9と同様に、系統群F内にある。
【0006】
クラッベ病(Krabbe disease)(グロボイド細胞白質ジストロフィー、GLD)は、加水分解酵素であるガラクトシルセラミダーゼ(GALC)をコードする遺伝子の変異によって引き起こされる常染色体劣性のリソソーム蓄積症(LSD)である(Wenger D.A.,et al.(2000)Mol Genet Metab.70(1):1-9)。この酵素は、ほぼ髄鞘のみに見られるガラクトシルセラミド(セラミド)およびガラクトシルスフィンゴシン(サイコシン)を含む特定のガラクト脂質の分解に関与している。クラッベ病では、GALC欠損症は、リソソーム内のサイコシン(ただし、ガラクトシルセラミドではない)の毒性蓄積を引き起こす(Svennerholm et al.,1980)。サイコシンの蓄積は、ミエリンを産生するCNSのオリゴデンドロサイトおよびPNSのシュワン細胞に対して、特に毒性があり、これらの細胞型の急速かつ広範な死をもたらす。CNSおよびPNSの両方におけるミエリン分解は、反応性アストロサイトのグリオーシスおよび多核巨マクロファージ(「グロボイド細胞」)の浸潤を伴う(Suzuki K.(2003)J Child Neurol.18(9):595-603)。ガラクトシルセラミドは、主に別の酵素であるGM1ガングリオシドβ-ガラクトシダーゼによる加水分解(Kobayashi T.,et al.(1985)J Biol Chem.260(28):14982-7)およびガラクトシルセラミド合成の停止に寄与するオリゴデンドロサイトの死(Svennerholm L.,et al.(1980)J Lipid Res.21(1):53-64)により、GALC活性が不在の場合は蓄積しない。
【0007】
現在、クラッベ病に対して利用可能な唯一の疾患修飾治療は、造血幹細胞移植(HSCT)であり、これは、多くの場合、臍帯血移植(UCBT)、同種末梢血幹細胞、または同種骨髄によって提供される。HSCTを使用した乳児性クラッベ病を有する患者の治療は、わずかに成功しただけであり、典型的には、1歳の誕生日を迎える前に症状を呈する。乳児性クラッベ病における明らかな症状の発症後に実施される場合、HSCTは、最小限の神経学的改善のみを提供し、実質的に生存率を改善しない(Escolar M.L.,et al.(2005)N Engl J Med.352(20):2069-81)。HSCTは、症候前患者で実施された場合に効果的であり得るが、それでも、運動転帰は悪い(Escolar M.L.,et al.(2005)N Engl J
Med.352(20):2069-81、Wright M.D.,et al.(2017)Neurology.89(13):1365-1372、van den Broek B.T.A.,et al.(2018)Blood Adv.2(1):49-60)。30日齢前に移植を受けた乳児は、後に移植を受ける乳児と比較して、より良好な生存率および機能的転帰を有した(Allewelt H.,et al.(2018)Biol Blood Marrow Transplant.24(11):2233-2238)。症候前の移植は、症状発症後の未治療のまたは治療された乳児性クラッベ病患者と比較して、顕著に良好な転帰をもたらすことが報告されており、進行性の中枢髄鞘形成、正常な受容言語、症状重症度の減衰、およびより長い生存期間を伴う(Escolar M.L.,et al.(2005)N Engl J Med.352(20):2069-81、Duffner P.K.,et al.(2009)Genet Med.11(6):450-4、Wright M.D.,et al.(2017)Neurology.89(13):1365-1372)。それでも、症状の出現前に治療されたほとんどの小児は、身長および体重について平均をかなり下回ったままであり、軽度の痙攣から独立した歩行の不能にわたって進行性の粗大運動遅延を有する(Escolar M.L.,et al.(2005)N Engl J Med.352(20):2069-81、Duffner P.K.,et al.(2009
)Genet Med.11(6):450-4)。一部の子供は、後天性小頭症、胃瘻造設の必要性、および構音障害を含む残存障害も有する(Duffner P.K.,et al.(2009)Genet Med.11(6):450-4)。さらに、HSCTは、CNS特異的疾患の病理にのみ影響を及ぼすように見える。末梢神経障害などのPNS病理に関連する臨床的特徴は、HSCTの影響を受けないままである。これらの結果は、HSCTの限界を強調するものであり、特に早期発症型では、疾患の急速な進行は、造血幹細胞が移植され、CNSに移行し、分化し、GALC分泌および交差補正(すなわち、矯正細胞によって分泌される酵素がGALC欠損細胞によって取り込まれるプロセス)を通して治療効果がもたらされるのに必要な時間を上回る。
【0008】
当該技術分野では、クラッベ病患者に対する改善された治療の必要性が、依然として存在する。
【発明の概要】
【0009】
組換えアデノ随伴ウイルス(rAAV)を含む組成物が提供され、中枢神経系の細胞を標的とするAAVカプシドと、(i)タンパク質の発現を指示する配列の制御下で、配列番号10のアミノ酸配列を有する成熟ガラクトシルセラミダーゼタンパク質をコードするガラクトシルセラミダーゼのコード配列、および(ii)ベクターゲノムをAAVカプシド中にパッケージングするために必要なAAV逆位末端反復を含むベクターゲノムと、を含み、ベクターゲノムが、AAVカプシド中にパッケージングされる。特定の実施形態では、AAVカプシドは、AAVhu68カプシドである。特定の実施形態では、コード配列は、配列番号9の核酸配列、またはそれと95%~99.9%同一の配列を有する。特定の実施形態では、コード配列は、配列番号10の成熟タンパク質、および中枢神経系のヒト細胞に好適な外因性シグナルペプチドをコードする。特定の実施形態では、調節配列は、ベータアクチンプロモーター、イントロン、および/またはウサギグロビンポリAを含む。特定の実施形態では、組成物は、ベクターゲノムCB7.CI.hGALC.rBGを有するrAAVを含む。
【0010】
特定の実施形態では、組換えアデノ随伴ウイルスが提供され、中枢神経系の細胞を標的とするAAVカプシドと、(i)成熟ガラクトシルセラミダーゼタンパク質の発現を指示する調節配列の制御下で、配列番号10のアミノ酸配列を有する成熟ガラクトシルセラミダーゼタンパク質をコードするガラクトシルセラミダーゼのコード配列、および(ii)ベクターゲノムをAAVカプシド中にパッケージングするために必要なAAV逆位末端配列を含むベクターゲノムと、を含む。特定の実施形態では、AAVカプシドは、AAVhu68カプシドである。特定の実施形態では、コード配列は、配列番号9の核酸配列、またはそれと95%~99.9%同一の配列を有する。特定の実施形態では、コード配列は、配列番号10の成熟タンパク質、および中枢神経系のヒト細胞に好適な外因性シグナルペプチドをコードする。特定の実施形態では、調節配列は、ベータアクチンプロモーター、イントロン、および/またはウサギグロビンポリAを含む。特定の実施形態では、ベクターゲノムは、CB7.CI.hGALC.RBGである。
(【0011】以降は省略されています)
この特許をJ-PlatPatで参照する
関連特許
他の特許を見る





 特許ウォッチ
特許ウォッチ